พลังงานที่ต้องส่งให้กับระบบเคมีเพิ่มเติมเพื่อ "เริ่มต้น" ปฏิกิริยาเรียกว่าพลังงานกระตุ้นสำหรับปฏิกิริยาที่กำหนดและทำหน้าที่เป็นสันพลังงานชนิดหนึ่งที่ต้องเอาชนะ.
ในปฏิกิริยาที่ไม่เร่งปฏิกิริยา แหล่งที่มาของพลังงานกระตุ้นคือการชนกันระหว่างโมเลกุล หากโมเลกุลที่ชนกันมีทิศทางที่เหมาะสมและการชนกันนั้นรุนแรงเพียงพอ ก็มีโอกาสที่พวกมันจะเกิดปฏิกิริยา
เป็นที่ชัดเจนว่าทำไมนักเคมีจึงใช้ขวดความร้อนเพื่อเร่งปฏิกิริยา: เมื่ออุณหภูมิสูงขึ้น ความเร็วของการเคลื่อนที่ด้วยความร้อนและความถี่ของการชนจะเพิ่มขึ้น แต่ภายใต้สภาวะของร่างกายมนุษย์คุณไม่สามารถให้ความร้อนแก่เซลล์ได้ซึ่งเป็นสิ่งที่ยอมรับไม่ได้ และปฏิกิริยาเกิดขึ้นและด้วยความเร็วที่ไม่สามารถบรรลุได้เมื่อทำในหลอดทดลอง สิ่งประดิษฐ์จากธรรมชาติอีกอย่างหนึ่งได้ผลที่นี่ - เอนไซม์ ซึ่งเราได้กล่าวถึงก่อนหน้านี้
ดังที่ได้กล่าวไปแล้ว ในระหว่างการเปลี่ยนแปลงทางเคมี ปฏิกิริยาเหล่านั้นสามารถเกิดขึ้นเองได้เอง โดยที่พลังงานที่มีอยู่ในผลิตภัณฑ์ของปฏิกิริยาน้อยกว่าในสารตั้งต้น ปฏิกิริยาอื่นๆ ต้องใช้พลังงานไหลเข้ามาจากภายนอก ปฏิกิริยาที่เกิดขึ้นเองสามารถเปรียบเทียบได้กับน้ำหนักที่ลดลง ในตอนแรก ภาระที่เหลือมีแนวโน้มที่จะตกลงมา ดังนั้นจึงลดพลังงานศักย์ลง
ในทำนองเดียวกัน ปฏิกิริยาที่เกิดขึ้นเมื่อเกิดขึ้นแล้วมีแนวโน้มที่จะก่อตัวเป็นสารที่มีพลังงานน้อยกว่า กระบวนการนี้ในระหว่างที่สามารถทำงานได้เรียกว่า โดยธรรมชาติ.
แต่ถ้าคุณเชื่อมต่อสิ่งของสองชิ้นด้วยวิธีใดวิธีหนึ่ง เมื่อตกลงมาอันที่หนักกว่าจะยกอันที่เบากว่าขึ้น และในกระบวนการทางเคมี โดยเฉพาะทางชีวเคมี ปฏิกิริยาที่ปล่อยพลังงานอาจทำให้เกิดปฏิกิริยาที่เกี่ยวข้องซึ่งต้องอาศัยการไหลเข้าของพลังงานจากภายนอก ปฏิกิริยาดังกล่าวเรียกว่า ผัน
ในสิ่งมีชีวิต ปฏิกิริยาคอนจูเกตเป็นเรื่องปกติมาก และการเกิดขึ้นของพวกมันเป็นตัวกำหนดปรากฏการณ์ที่ละเอียดอ่อนทั้งหมดที่มาพร้อมกับชีวิตและจิตสำนึก “ภาระหนัก” ที่ตกลงมาจะทำให้อีกอันหนึ่งที่เบากว่าเพิ่มขึ้นแต่ในปริมาณที่น้อยกว่า เมื่อเรารับประทานอาหารเราจะดูดซับสารที่มีพลังงานคุณภาพสูงจากดวงอาทิตย์ซึ่งจะสลายตัวในร่างกายและถูกปล่อยออกมาในที่สุด แต่ในขณะเดียวกันก็สามารถปล่อยพลังงานออกมาได้ในปริมาณที่เพียงพอต่อกระบวนการที่เรียกว่าชีวิต .
ในเซลล์ ตัวกลางพลังงานหลัก ซึ่งก็คือ "วงล้อขับเคลื่อน" ของชีวิต อะดีโนซีน ไตรฟอสเฟต (ATP) . ทำไมการเชื่อมต่อนี้ถึงน่าสนใจ? จากมุมมองทางชีวเคมี ATP เป็นโมเลกุลขนาดกลางที่สามารถเกาะหรือ "ปล่อย" กลุ่มฟอสเฟตปลายซึ่งอะตอมฟอสฟอรัสถูกล้อมรอบด้วยอะตอมออกซิเจน
การก่อตัวของ ATP เกิดขึ้นจากอะดีโนซีนไดฟอสเฟต (ADP) เนื่องจากพลังงานที่ปล่อยออกมาระหว่างการออกซิเดชันทางชีวภาพของกลูโคส ในทางกลับกัน การทำลายพันธะฟอสเฟตใน ATP จะปล่อยพลังงานจำนวนมาก พันธะดังกล่าวเรียกว่าพลังงานสูงหรือพลังงานสูง โมเลกุล ATP ประกอบด้วยพันธะดังกล่าว 2 พันธะ ซึ่งการไฮโดรไลซิสจะปล่อยพลังงานออกมาเทียบเท่ากับ 12-14 กิโลแคลอรี
ไม่มีใครรู้ว่าทำไมในกระบวนการวิวัฒนาการธรรมชาติจึง "เลือก" ATP เป็นสกุลเงินพลังงานของเซลล์ แต่สามารถสันนิษฐานได้ด้วยเหตุผลหลายประการ ในทางอุณหพลศาสตร์ โมเลกุลนี้ค่อนข้างไม่เสถียร ดังที่เห็นได้จากพลังงานจำนวนมากที่ปล่อยออกมาในระหว่างการไฮโดรไลซิส
แต่ในขณะเดียวกันอัตราการไฮโดรไลซิสของเอนไซม์ของ ATP ภายใต้สภาวะปกติจะต่ำมาก กล่าวคือ โมเลกุล ATP มีความเสถียรทางเคมีสูง ทำให้กักเก็บพลังงานได้อย่างมีประสิทธิภาพ
โมเลกุล ATP ขนาดที่เล็กทำให้สามารถแพร่กระจายไปยังส่วนต่างๆ ของเซลล์ได้อย่างง่ายดาย ซึ่งจำเป็นต้องใช้พลังงานในการทำงานใดๆ ในที่สุด ATP ครองตำแหน่งระดับกลางในระดับของสารประกอบพลังงานสูง ซึ่งทำให้มีความสามารถรอบด้าน ช่วยให้สามารถถ่ายโอนพลังงานจากสารประกอบพลังงานสูงไปยังสารประกอบพลังงานต่ำได้
ดังนั้น ATP จึงเป็นรูปแบบสากลหลักในการกักเก็บพลังงานของเซลล์ซึ่งเป็นเชื้อเพลิงของเซลล์ที่พร้อมใช้ได้ตลอดเวลา และผู้จัดหาพลังงานหลักให้กับเซลล์ดังที่เราได้กล่าวไปแล้วก็คือ กลูโคส ที่ได้จากการสลายคาร์โบไฮเดรต “การเผาไหม้” ในร่างกาย กลูโคสจะก่อให้เกิดคาร์บอนไดออกไซด์และน้ำ และกระบวนการนี้ทำให้เกิดปฏิกิริยาการหายใจและการย่อยอาหารของเซลล์ คำว่า “ไหม้” ในกรณีนี้คือรูปภาพ ไม่มีเปลวไฟภายในร่างกาย และพลังงานถูกสกัดด้วยวิธีการทางเคมีหลายขั้นตอน
ในระยะแรก เกิดขึ้นในไซโตพลาสซึมโดยไม่มีออกซิเจน โมเลกุลของกลูโคสจะแบ่งออกเป็นสองส่วน (กรดไพรูวิกสองโมเลกุล) และระยะนี้เรียกว่า ไกลโคไลซิส . สิ่งนี้จะปล่อยพลังงาน 50 กิโลแคลอรี/โมล (นั่นคือ 7% ของพลังงานที่มีอยู่ในกลูโคส) ซึ่งส่วนหนึ่งจะสลายไปเป็นความร้อน และอีกส่วนหนึ่งถูกใช้ไปกับการก่อตัวของโมเลกุล ATP สองตัว
การสกัดพลังงานจากกลูโคสในภายหลังส่วนใหญ่เกิดขึ้นในไมโตคอนเดรียซึ่งเป็นสถานีพลังงานของเซลล์ซึ่งสามารถเปรียบเทียบงานกับเซลล์กัลวานิกได้ ในแต่ละขั้นตอน อิเล็กตรอนและไฮโดรเจนไอออนจะถูกกำจัดออก และในที่สุดกลูโคสก็สลายตัวเป็นคาร์บอนไดออกไซด์และน้ำ
ใน ไมโตคอนเดรีย อิเล็กตรอนและไฮโดรเจนไอออนถูกใส่เข้าไปในเอนไซม์รีดอกซ์สายโซ่เดี่ยว (สายโซ่ทางเดินหายใจ) โดยผ่านจากตัวกลางไปยังตัวกลางจนกระทั่งพวกมันรวมตัวกับออกซิเจน และในขั้นตอนนี้ ไม่ใช่ออกซิเจนในอากาศที่ใช้สำหรับการเกิดออกซิเดชัน แต่เป็นออกซิเจนจากน้ำและกรดอะซิติก
ออกซิเจนในอากาศเป็นตัวรับไฮโดรเจนตัวสุดท้าย ซึ่งทำให้กระบวนการหายใจของเซลล์เสร็จสิ้น ซึ่งเป็นเหตุผลว่าทำไมจึงจำเป็นต่อชีวิต ดังที่ทราบกันดีว่าปฏิกิริยาของก๊าซออกซิเจนและไฮโดรเจนนั้นมาพร้อมกับการระเบิด (ปล่อยพลังงานจำนวนมากทันที)
สิ่งนี้ไม่ได้เกิดขึ้นในสิ่งมีชีวิตเนื่องจากไม่ได้เกิดก๊าซไฮโดรเจน และเมื่อถึงเวลาที่มันจับกับออกซิเจนในอากาศ ปริมาณพลังงานอิสระจะลดลงมากจนปฏิกิริยาการก่อตัวของน้ำดำเนินไปอย่างสงบอย่างสมบูรณ์ (ดู ภาพที่ 1).
กลูโคสเป็นสารตั้งต้นหลัก แต่ไม่ใช่สารตั้งต้นเพียงอย่างเดียวสำหรับการผลิตพลังงานในเซลล์ นอกจากคาร์โบไฮเดรตแล้ว ร่างกายของเรายังได้รับไขมัน โปรตีน และสารอื่นๆ จากอาหาร ซึ่งหลังจากการสลายแล้วยังเป็นแหล่งพลังงานอีกด้วย ซึ่งกลายเป็นสารที่รวมอยู่ในปฏิกิริยาทางชีวเคมีที่เกิดขึ้นในเซลล์
การวิจัยพื้นฐานในสาขาทฤษฎีสารสนเทศนำไปสู่การเกิดขึ้นของแนวคิดนี้ พลังงานสารสนเทศ (หรือพลังงานของผลกระทบของข้อมูล) เป็นความแตกต่างระหว่างความแน่นอนและความไม่แน่นอน ในที่นี้ ฉันอยากจะทราบว่าเซลล์ใช้และใช้พลังงานข้อมูลเพื่อขจัดความไม่แน่นอนในทุกช่วงเวลาของวงจรชีวิต สิ่งนี้นำไปสู่การดำเนินวงจรชีวิตโดยไม่เพิ่มเอนโทรปี
การหยุดชะงักของกระบวนการเผาผลาญพลังงานภายใต้อิทธิพลของอิทธิพลต่าง ๆ นำไปสู่ความล้มเหลวในแต่ละขั้นตอนและผลจากความล้มเหลวเหล่านี้ทำให้เกิดการหยุดชะงักของระบบย่อยของกิจกรรมชีวิตของเซลล์และสิ่งมีชีวิตทั้งหมดโดยรวม หากจำนวนและความชุกของความผิดปกติเหล่านี้เกินความสามารถในการชดเชยของกลไกสภาวะสมดุลในร่างกาย ระบบก็จะออกจากการควบคุมและเซลล์จะหยุดทำงานพร้อมกัน ในระดับร่างกายสิ่งนี้จะแสดงออกมาในรูปแบบของสภาวะทางพยาธิวิทยาต่างๆ
ดังนั้นการขาดวิตามินบี 1 ซึ่งเกี่ยวข้องกับการทำงานของเอนไซม์บางชนิดนำไปสู่การปิดกั้นการเกิดออกซิเดชันของกรดไพรูวิก ฮอร์โมนไทรอยด์ส่วนเกินขัดขวางการสังเคราะห์ ATP เป็นต้น การเสียชีวิตจากกล้ามเนื้อหัวใจตาย พิษจากคาร์บอนมอนอกไซด์ หรือพิษจากโพแทสเซียมไซยาไนด์ ยังเกี่ยวข้องกับการปิดกั้นกระบวนการหายใจของเซลล์โดยการยับยั้งหรือแยกปฏิกิริยาตามลำดับออก การออกฤทธิ์ของสารพิษจากแบคทีเรียหลายชนิดเป็นไปโดยอ้อมผ่านกลไกที่คล้ายกัน
ดังนั้นการทำงานของเซลล์ เนื้อเยื่อ อวัยวะ ระบบอวัยวะ หรือสิ่งมีชีวิตในฐานะระบบนั้นได้รับการสนับสนุนโดยกลไกการควบคุมตนเอง ซึ่งแนวทางที่เหมาะสมที่สุดจะได้รับการรับประกันโดยกระบวนการทางชีวฟิสิกส์ ชีวเคมี พลังงานและข้อมูล
ชุดของค่าทั้งหมดของพารามิเตอร์ทางอุณหพลศาสตร์ที่จำเป็นในการอธิบายระบบเรียกว่า สถานะทางอุณหพลศาสตร์ .
ลักษณะทางกายภาพของระบบการเปลี่ยนแปลงที่เมื่อระบบเปลี่ยนจากสถานะหนึ่งไปอีกสถานะหนึ่งจะถูกกำหนดโดยค่าของพารามิเตอร์ของสถานะเริ่มต้นและขั้นสุดท้ายและไม่ขึ้นอยู่กับการเปลี่ยนแปลงเรียกว่า ฟังก์ชั่นของรัฐ (ศักยภาพทางอุณหพลศาสตร์).
หน้าที่ของรัฐคือ:
· กำลังภายใน;
· เอนทาลปี;
· เอนโทรปี;
· พลังงานฟรี
· ศักยภาพทางเคมีและไฟฟ้าเคมี
เรียกว่าปริมาณของปริมาณใด ๆ ที่ถ่ายโอนต่อหน่วยเวลาผ่านพื้นผิวหนึ่ง ๆ ไหล ค่านี้
ปรากฏการณ์ที่กระบวนการหนึ่งทำให้แน่ใจว่ากระบวนการที่สองจะเกิดขึ้นอย่างกระฉับกระเฉงเรียกว่า การจับคู่ .
กระบวนการสร้างพลังงานเรียกว่า...
การผันคำกริยา. กระบวนการที่ใช้พลังงานเรียกว่า ผัน
.
กฎข้อที่หนึ่งและสองของอุณหพลศาสตร์ตามกฎข้อที่หนึ่งของอุณหพลศาสตร์ซึ่งอ่านได้ดังนี้: ความร้อนที่มอบให้กับระบบนั้นถูกใช้ไปในการเพิ่มพลังงานภายในของระบบและระบบที่ทำงานด้วยแรงภายนอก พลังงานประเภทต่าง ๆ สามารถแปลงร่างเป็นกันและกันได้ แต่ในระหว่าง การเปลี่ยนแปลงเหล่านี้พลังงานจะไม่หายไปและไม่ปรากฏขึ้นจากความว่างเปล่า ซึ่งหมายความว่าสำหรับระบบปิด
∆U = ∆Q –W โดยที่ ∆U คือการเปลี่ยนแปลงพลังงานภายในของระบบ ∆Q คือความร้อนที่ระบบดูดซับ W คืองานที่ระบบทำ [พลังงานภายในแตกต่างจากความร้อน และการทำงานจะเปลี่ยนแปลงเหมือนเดิมเสมอเมื่อเปลี่ยนจากสถานะหนึ่งไปอีกสถานะหนึ่ง โดยไม่คำนึงถึงเส้นทางของการเปลี่ยนแปลง!]
การเปลี่ยนแปลงพลังงานความร้อน ∆Q ของระบบแยกเดี่ยวจะเป็นสัดส่วนกับอุณหภูมิสัมบูรณ์ T ซึ่งก็คือ ∆Q = T ∙ ∆S โดยที่ ∆S คือสัมประสิทธิ์สัดส่วนที่เรียกว่าการเปลี่ยนแปลงในเอนโทรปี
กฎข้อที่สองของอุณหพลศาสตร์มีอยู่ในสองสูตร สูตรแรก (สูตรของคลอเซียส) มีดังต่อไปนี้ การถ่ายเทความร้อนจากวัตถุที่มีอุณหภูมิต่ำกว่าไปยังวัตถุที่มีอุณหภูมิสูงกว่านั้นเป็นไปไม่ได้ สูตรที่สอง (สูตรของทอมสัน) บอกว่าเป็นไปไม่ได้ที่จะสร้างเครื่องจักรการเคลื่อนที่ตลอดเวลาประเภทนี้ นั่นคือ กระบวนการแบบวนรอบซึ่งเป็นผลมาจากการที่ความร้อนทั้งหมดที่ระบบดูดซับไว้จะถูกนำไปใช้ในการทำงาน ตามกฎข้อที่สองของอุณหพลศาสตร์ เอนโทรปีของระบบแยกจะเพิ่มขึ้นในกระบวนการที่ไม่สามารถย้อนกลับได้ แต่ยังคงไม่เปลี่ยนแปลงในกระบวนการที่ย้อนกลับได้ เอนโทรปีเป็นฟังก์ชันของสถานะของระบบ ส่วนต่างของกระบวนการที่ผันกลับได้เล็กน้อยจะเท่ากับอัตราส่วนของปริมาณความร้อนเพียงเล็กน้อยที่ส่งให้กับระบบต่ออุณหภูมิสัมบูรณ์ของกระบวนการหลัง (ΔS=ΔQ:T) หน่วยของเอนโทรปีคือ J/K เอนโทรปีเป็นการวัดความผิดปกติของระบบ หากเอนโทรปีเพิ่มขึ้น หมายความว่าระบบมีแนวโน้มที่จะเคลื่อนเข้าสู่สถานะที่มีความน่าจะเป็นทางอุณหพลศาสตร์สูงกว่า กล่าวคือ เข้าสู่สถานะที่มีลำดับน้อยลง ข้อสรุปตามมาจากกฎข้อที่สองของอุณหพลศาสตร์: ที่อุณหภูมิคงที่ พลังงานความร้อนไม่สามารถแปลงเป็นงานเครื่องกลได้ เนื่องจากพลังงานความร้อนเกิดจากการเคลื่อนที่ของอนุภาคอย่างวุ่นวาย ผลรวมของเวกเตอร์ความเร็วของอนุภาคเหล่านี้ในทิศทางใดๆ จึงเป็นศูนย์ เฉพาะพลังงานที่แสดงถึงการเคลื่อนที่ในทิศทางเดียวของร่างกาย (พลังงานจลน์ของวัตถุที่บินได้ พลังงานของไอออนที่เคลื่อนที่หรืออิเล็กตรอนในสนามไฟฟ้า) เท่านั้นที่สามารถแปลงเป็นงานเชิงกลได้
ข้อสรุปตามกฎหมายสองฉบับ:
กฎข้อแรกกำหนดความสัมพันธ์เชิงปริมาณระหว่างความร้อน งาน และการเปลี่ยนแปลงพลังงานภายใน แต่ไม่ได้กำหนดทิศทางของกระบวนการทางอุณหพลศาสตร์ จะดำเนินการอยู่เสมอและสำหรับระบบใดๆ ความสัมพันธ์พื้นฐานของอุณหพลศาสตร์: TΔS ≥ ΔU+W
กฎข้อที่สองเป็นกฎทางสถิติและใช้ได้กับระบบที่มีอนุภาคจำนวนมากและมีจำนวนจำกัด มันบ่งบอกถึงทิศทางที่เป็นไปได้มากที่สุดของกระบวนการ หากมีการระบุว่ากระบวนการนี้เป็นไปไม่ได้ ก็ควรเข้าใจว่าความน่าจะเป็นที่จะเกิดขึ้นนั้นมีอยู่ แต่จะน้อยมาก
ตารางที่ 1. ศักย์ทางอุณหพลศาสตร์
การเปลี่ยนแปลงพลังงานในเซลล์ของสิ่งมีชีวิตในเซลล์ที่มีชีวิต พลังงานเคมีซึ่งเก็บไว้ในสารประกอบอินทรีย์จะถูกแปลงเป็นพลังงานออสโมติก พลังงานไฟฟ้า และพลังงานกล ตัวอย่างเช่น พลังงานเคมีของกลูโคสจะถูกแปลงในระหว่างการออกซิเดชันของเซลล์ ส่วนหนึ่งเป็นความร้อน และอีกส่วนหนึ่งเป็นพลังงานของพันธะมหภาคของ ATP เนื่องจากการไฮโดรไลซิสของ ATP การถ่ายโอนของสารจากบริเวณที่ต่ำกว่าไปยังบริเวณที่มีความเข้มข้นสูงกว่าสามารถเกิดขึ้นได้ (งานออสโมติก) การถ่ายโอนไอออนไปยังพื้นที่ที่มีศักยภาพทางไฟฟ้าสูงกว่า (งานไฟฟ้า) และในร่างกายของสัตว์ - การหดตัวของกล้ามเนื้อ (งานเครื่องกล) ในกรณีนี้ พลังงานเคมีส่วนหนึ่งของ ATP จะถูกแปลงเป็นพลังงานออสโมติก พลังงานไฟฟ้า และพลังงานกล
พลังงานฟรีและศักย์ไฟฟ้าเคมีพลังงานไฟฟ้า ออสโมติก และเคมีของเซลล์ถูกใช้ในการทำงาน กล่าวคือ สำหรับการเคลื่อนที่ของอนุภาคโดยตรงต่อแรงที่กระทำต่อพวกมัน การวัดเชิงปริมาณของการเปลี่ยนแปลงของพลังงานประเภทนี้คือการเปลี่ยนแปลงของพลังงานอิสระ (∆F) ΔF คือพลังงานอิสระของเฮล์มโฮลทซ์ (ΔF =ΔU – TΔS) เนื่องจากขึ้นอยู่กับเงื่อนไขของกระบวนการ โดยเฉพาะอย่างยิ่งความเข้มข้นของสารที่ทำปฏิกิริยา พวกเขาจึงเริ่มใช้สิ่งที่เรียกว่าศักย์ทางอุณหพลศาสตร์กิ๊บส์ที่ 1 โมลของสาร ΔG ในวิชาเคมีสำหรับอนุภาคที่ไม่มีประจุจะเรียกว่าศักย์เคมี - μสำหรับอนุภาคที่มีประจุ - ศักย์ไฟฟ้าเคมี - μ
การเกิดปฏิกิริยาเคมีในสถานะของเหลวไม่ได้เปลี่ยนความดัน แต่สามารถเปลี่ยนปริมาตรได้ ดังนั้น สำหรับระบบดังกล่าว แทนที่จะเปลี่ยนพลังงานภายใน ระบบจะใช้การเปลี่ยนแปลงของเอนทัลปี (∆H) ซึ่งเท่ากับ ∆U+p∆V โดยที่ p คือความดัน ∆V คือการเปลี่ยนแปลงของปริมาตร [หมายเหตุ: เอนทาลปีเป็นฟังก์ชันของสถานะของระบบเทอร์โมไดนามิกส์ที่มีพารามิเตอร์อิสระของเอนโทรปีและความดัน] ตามกฎของอุณหพลศาสตร์ มีความสัมพันธ์ระหว่างการเปลี่ยนแปลงของพลังงานภายในและการเปลี่ยนแปลงของเอนทาลปี: ∆G = ∆H -T∆S (ที่ t และ p = const) โดยที่ ΔG คือศักย์ทางอุณหพลศาสตร์ของ Gibbs, ΔH คือพลังงานภายใน T * ΔS คือพลังงานความร้อน
ในระบบเคมีกายภาพ การเปลี่ยนแปลงของพลังงานอิสระมักจะอธิบายผ่านการเปลี่ยนแปลงศักย์เคมีไฟฟ้า (∆μ): ∆G=m∙∆μ โดยที่ m คือปริมาณของสาร (โมล) ในระบบ การเปลี่ยนแปลงศักย์ไฟฟ้าเคมีระหว่างการเปลี่ยนระบบจากสถานะ 1 เป็นสถานะ 2 ถูกกำหนดโดยการเปลี่ยนแปลงของพลังงานเคมี ออสโมติก และไฟฟ้า: ∆μ = μ 02 -μ 01 +RT ln (c 2 /c 1) + zF (φ 2 -φ 1) จากนั้น ∆G = m μ 02 -μ 01 +RT ln (c 2 /c 1) + zF (φ 2 -φ 1)
ความหมายทางกายภาพของศักย์ไฟฟ้าเคมีคือการเปลี่ยนแปลงเท่ากับงานที่ต้องใช้เพื่อ:
1. สังเคราะห์สาร 1 โมล (สถานะ 2) จากสารเริ่มต้น (สถานะ 1) และวางไว้ในตัวทำละลาย (คำสั่งμ 02 -μ 01) - งานเคมี
2. ทำให้สารละลายเข้มข้นจากความเข้มข้นตั้งแต่ 1 ถึง c 2 [เทอม RT ln (c 2 /c 1)] – งานออสโมติก
3. เอาชนะแรงผลักไฟฟ้าที่เกิดขึ้นเมื่อมีความต่างศักย์ (φ 2 -φ 1) ระหว่างสารละลาย [คำว่า zF (φ 2 -φ 1] – งานไฟฟ้า
ควรสังเกตว่าเงื่อนไขสามารถเป็นได้ทั้งเชิงบวกและเชิงลบ
กฎข้อที่สองของอุณหพลศาสตร์และสภาวะสมดุลกฎข้อที่สองของอุณหพลศาสตร์ระบุว่าพลังงานอิสระไม่สามารถเพิ่มขึ้นในระบบที่แยกได้ กล่าวอีกนัยหนึ่ง ในระบบที่ ∆H = 0, ∆G = -T∆S ≤0 ตราบใดที่การเปลี่ยนแปลงพลังงานในระบบที่กำหนดจะมาพร้อมกับการเปลี่ยนพลังงานประเภทต่างๆ เข้าหากันโดยไม่เปลี่ยนเป็นความร้อน นั่นคือ ∆G=0 กระบวนการทั้งหมดนี้สามารถย้อนกลับได้ แต่ทันทีที่พลังงานส่วนหนึ่งกลายเป็นความร้อน กระบวนการนี้ก็จะกลับคืนไม่ได้ แนวคิดเรื่องการพลิกกลับของกระบวนการมีความเกี่ยวข้องกับแนวคิดเรื่องสมดุลแบบไดนามิก สมดุลคือสถานะของระบบที่แต่ละอนุภาคสามารถเคลื่อนที่จากบางสถานะ 1 ไปยังสถานะ 2 และย้อนกลับได้ แต่โดยทั่วไปแล้วสัดส่วนของสถานะ 1 และสถานะ 2 ในระบบจะไม่เปลี่ยนแปลง ในระบบเคมีกายภาพ กระบวนการอยู่ในสมดุลโดยที่ ∆μ = ∆G/m = 0 นั่นคือ μ 02 -μ 01 +RT ln (c 2 /c 1) + zF (φ 2 -φ 1) = 0
สารตั้งต้นและผลิตภัณฑ์จากปฏิกิริยาทางชีวเคมีหรือไอออนทั้งสองด้านของเมมเบรนอาจอยู่ในสภาวะสมดุล ดังนั้นจึงมีการประยุกต์สมการที่อธิบายสถานะสมดุลของระบบ:
1. สมการของค่าคงที่สมดุลเคมี: ∆μ 0 = -RT lnK โดยที่ K คือค่าคงที่สมดุล
2. สมการของศักยภาพของเมมเบรนสมดุล (สมการ Nernst): หากเยื่อหุ้มเซลล์สามารถซึมผ่านไปยังไอออนใดไอออนหนึ่งได้ ศักยภาพของเมมเบรนสมดุลจะถูกสร้างขึ้นบนเมมเบรน: φ M = φ 1 – φ 2 = RT/zF lnc 1 / c 2 ที่อุณหภูมิ 37С 0 φ Μ = 60 ln (s 1 / s 2) mV เพื่อการเขียนที่กระชับยิ่งขึ้น เราได้แนะนำแนวคิดเกี่ยวกับศักยภาพไร้มิติ ψ Μ ซึ่งเท่ากับ ln(c 1 /c 2) จากนั้นสมการ Nernst จะมีหน้าตาดังนี้: ψ Μ = ψ 1 – ψ 2 = ln(c 1 /ค2)
3. การกระจายของ Boltzmann: หากในโมเลกุลมีระดับพลังงานอิเล็กทรอนิกส์สองระดับที่มีพลังงาน E 1 และ E 2 ดังนั้นประชากรของระดับเหล่านี้ที่มีอิเล็กตรอนอยู่ในสภาวะสมดุลสามารถพบได้: ∆E = E 2 – E 1
การหาค่าพารามิเตอร์ทางอุณหพลศาสตร์ของระบบชีวภาพโดยการทดลองในการกำหนดพารามิเตอร์ทางอุณหพลศาสตร์ของระบบชีวภาพ มีการใช้สองวิธี: การหาปริมาณความร้อน (แคลอรี่) และการวัดค่าคงที่สมดุล เนื่องจากวัตถุที่อยู่ในแคลอริมิเตอร์ไม่ก่อให้เกิดงาน การเปลี่ยนแปลงของพลังงาน (เอนทาลปี) จึงถือได้เท่ากับปริมาณความร้อนที่ปล่อยออกมา ∆Q นี่เป็นวิธีที่พบการเปลี่ยนแปลงของเอนทัลปี ∆H ในระหว่างการศึกษากระบวนการทางชีวฟิสิกส์หรือปฏิกิริยาทางชีวเคมี อีกวิธีหนึ่งในการศึกษาพารามิเตอร์ทางอุณหพลศาสตร์คือการวัดค่าคงที่สมดุลที่อุณหภูมิต่างๆ แต่วิธีนี้ใช้ได้เฉพาะเมื่อการเปลี่ยนแปลงเอนทาลปีและการเปลี่ยนแปลงเอนโทรปีไม่ได้ขึ้นอยู่กับอุณหภูมิ ในกรณีนี้ จะใช้สมการแวนต์ ฮอฟฟ์: lnK = -∆H/RT + ∆S/R (สำหรับสารหนึ่งโมล)
สิ่งมีชีวิตในฐานะระบบอุณหพลศาสตร์เมื่อใช้อุณหพลศาสตร์กับระบบชีวภาพจำเป็นต้องคำนึงถึงลักษณะเฉพาะของระบบสิ่งมีชีวิตด้วย:
1) ระบบชีวภาพเปิดให้สสารและพลังงานไหลผ่าน
2) กระบวนการในระบบสิ่งมีชีวิตไม่สามารถย้อนกลับได้
3) ระบบสิ่งมีชีวิตยังห่างไกลจากความสมดุล
4) ระบบทางชีววิทยาเป็นแบบเฮเทอโรเฟสิก มีโครงสร้าง และแต่ละเฟสอาจมีโมเลกุลจำนวนน้อย
ทั้งหมดนี้ทำให้ระบบทางชีววิทยาแตกต่างจากระบบแยกเดี่ยวที่ใกล้สมดุล ดังนั้น เพื่อที่จะอธิบายคุณสมบัติของระบบสิ่งมีชีวิตได้เพียงพอมากขึ้น จึงจำเป็นต้องใช้อุณหพลศาสตร์ของกระบวนการที่ไม่สามารถย้อนกลับได้ อุณหพลศาสตร์ของกระบวนการที่ไม่สามารถย้อนกลับได้จะแตกต่างจากอุณหพลศาสตร์แบบคลาสสิก โดยพิจารณาถึงวิถีทางของกระบวนการเมื่อเวลาผ่านไป แนวคิดพื้นฐานในอุณหพลศาสตร์คลาสสิกคือแนวคิดเรื่องสภาวะสมดุล ในอุณหพลศาสตร์ของกระบวนการที่ไม่สามารถย้อนกลับได้ แนวคิดที่สำคัญคือแนวคิดเกี่ยวกับสถานะคงที่ของระบบ
หมายเหตุ: ต้องคำนึงว่าสิ่งมีชีวิตมีการพัฒนาและเปลี่ยนแปลงอยู่ตลอดเวลา ดังนั้นโดยรวมแล้ว ไม่ใช่ระบบที่อยู่กับที่ ในกรณีนี้มีการรับสมัคร: ในช่วงเวลาสั้น ๆ สถานะของบางส่วนจะได้รับการยอมรับว่านิ่ง
ตรงกันข้ามกับสมดุลทางอุณหพลศาสตร์ สถานะคงที่มีลักษณะเฉพาะ
· การไหลเข้าของสารเข้าสู่ระบบอย่างต่อเนื่องและการกำจัดผลิตภัณฑ์เมตาบอลิซึม
·ค่าใช้จ่ายคงที่ของพลังงานอิสระซึ่งรักษาความคงที่ของความเข้มข้นของสารในระบบ
· ความคงที่ของพารามิเตอร์ทางอุณหพลศาสตร์ (รวมถึงพลังงานภายในและเอนโทรปี)
ระบบที่อยู่ในสถานะคงที่สามารถเป็นได้ทั้งแบบปิดหรือเปิด ระบบเปิดสามารถดำรงอยู่ได้ก็ต่อเมื่อพลังงานไหลเข้ามาจากภายนอกและพลังงานไหลออกสู่สิ่งแวดล้อม ในระบบชีวภาพ การไหลที่สำคัญที่สุดคือการไหลของสารและประจุไฟฟ้า
การไหลของสารอันเป็นผลมาจากการแพร่กระจายและการแพร่กระจายด้วยไฟฟ้า 1.แรงผลักดันหลักในการเคลื่อนย้ายอนุภาคโดยการแพร่กระจายอย่างง่ายคือการไล่ระดับความเข้มข้น การไหลของสารอันเป็นผลจากการแพร่กระจายผ่านเยื่อหุ้มเซลล์คำนวณตามกฎของ Fick สำหรับการถ่ายโอนสารแบบพาสซีฟผ่านเมมเบรน: Φ = –DK/l (c in -c in) = –P (c in - c in) โดยที่ Φ คือการไหล; D – ค่าสัมประสิทธิ์การแพร่กระจาย; K คือสัมประสิทธิ์การกระจายของสารระหว่างเมมเบรนและเฟสน้ำโดยรอบ ล. – ความหนาของเมมเบรน; c cc – ความเข้มข้นของอนุภาคภายในเซลล์ с in – ความเข้มข้นของอนุภาคนอกเซลล์ P – ค่าสัมประสิทธิ์การซึมผ่าน หากเราพิจารณาการแพร่กระจายจากมุมมองของการแปลงพลังงาน การคำนวณจะต้องดำเนินการโดยใช้สมการต่อไปนี้: Φ = – uc (dG/dx) โดยที่ u = D/RT คือสัมประสิทธิ์สัดส่วนที่ขึ้นอยู่กับอัตราการแพร่ ของโมเลกุลและเรียกว่าการเคลื่อนไหว ดังนั้นการไหลจึงเป็นสัดส่วนกับความเข้มข้นของสารและการไล่ระดับของศักย์ทางอุณหพลศาสตร์ในทิศทางของการไหล
2. แรงผลักดันหลักในการถ่ายโอนอนุภาคที่มีประจุในกรณีที่ไม่มีการไล่ระดับความเข้มข้นคือสนามไฟฟ้า ในกรณีนี้ จะใช้สมการทฤษฎีบท: Φ = – cu (dμ/dx) โดยที่ μ คือศักย์เคมีไฟฟ้า ดังนั้น ฟลักซ์จะเท่ากับผลคูณของความเข้มข้นของตัวพาโดยการเคลื่อนที่และการไล่ระดับของศักย์ไฟฟ้าเคมี เครื่องหมาย “–” บ่งบอกว่ากระแสไหลไปในทิศทางที่ลดลง μ นอกจากนี้ ยังใช้สมการการแพร่กระจายคลื่นไฟฟ้าของเนิร์นสต์–พลังค์ด้วย: Φ = –uRT (dc/dx) –cuz Fdφ/dx
การไหลและแรงทางอุณหพลศาสตร์ที่กำหนดการเกิดขึ้นของกระบวนการสำคัญแสดงไว้ในตารางที่ 3
ตารางที่ 3 การผันการไหลและแรงในอุณหพลศาสตร์ที่ไม่สมดุล
อุณหพลศาสตร์ของสถานะคงตัวระบบเปิดมีคุณสมบัติเฉพาะ: การควบรวมของกระแสและการเกิดขึ้นของสถานะคงที่ คุณลักษณะของระบบเปิดเหล่านี้อธิบายได้ด้วยอุณหพลศาสตร์ของกระบวนการเชิงเส้นที่ไม่สามารถย้อนกลับได้ อธิบายการเกิดขึ้นพร้อมๆ กันของกระบวนการนิ่งต่างๆ ที่สัมพันธ์กัน Onsager ได้กำหนดทฤษฎีอุณหพลศาสตร์ของกระบวนการเชิงเส้นที่ไม่สามารถย้อนกลับได้ พื้นฐานการทดลองของทฤษฎีนี้คือกฎปรากฏการณ์วิทยาที่สร้างความสัมพันธ์เชิงเส้นตรงระหว่างกระแสและแรงที่ทำให้เกิดกระแส (ดูตารางที่ 2) สมมติว่ามีการไหลสองแบบในระบบ - การไหลของความร้อน (Φ 1) และการไหลของมวลการแพร่กระจาย (Φ 2) และแรงทั่วไปสองแบบ - ความแตกต่างของอุณหภูมิ X 1 และความแตกต่างของความเข้มข้น X 2 ตามข้อมูลของ Onsager ในระบบเปิด แต่ละการไหลขึ้นอยู่กับแรงทั้งหมดที่มีอยู่ และในทางกลับกัน นั่นคือ
Φ 1 = ยาว 11 X 1 + ยาว 12 X 2
Φ 2 = ยาว 21 X 1 + ยาว 22 X 2
โดยที่ L 12 และอื่นๆ เป็นสัมประสิทธิ์สัดส่วนระหว่างการไหล 1 และแรง 2 เป็นต้น
สมการเหล่านี้เรียกว่าสมการเชิงปรากฏการณ์วิทยา สิ่งเหล่านี้บ่งบอกถึงการพึ่งพาการไหลของอินพุตและเอาท์พุตของแรงคอนจูเกตและแรงที่ไม่ใช่คอนจูเกต ดังที่ Onsager แสดงให้เห็น ใกล้สมดุลแล้วสัมประสิทธิ์สัดส่วนระหว่างกระแสจะเท่ากัน (L 12 = L 21) กล่าวอีกนัยหนึ่ง การกระทำที่เท่าเทียมกันทำให้เกิดการตอบสนองที่เท่าเทียมกัน ตัวอย่างเช่น ผลการเบรกที่ตัวทำละลายที่กำลังเคลื่อนที่มีต่อตัวถูกละลายจะเท่ากับความต้านทานที่ตัวถูกละลายกระทำต่อตัวทำละลาย
ในธรรมชาติ มีสถานการณ์หนึ่งที่กระแสที่มาพร้อมกับพลังงานที่เพิ่มขึ้นไม่สามารถไหลได้อย่างอิสระ แต่สามารถไหลภายใต้การกระทำของแรงบางอย่างได้ ปรากฏการณ์นี้เรียกว่าการผันคำกริยา เกณฑ์สำหรับความเป็นไปได้ของการผันกระแสในระบบคือค่าบวกของฟังก์ชันการกระจาย ψ = Τ/V dS/dt ≥ 0 โดยที่ Τ คืออุณหภูมิสัมบูรณ์ dS/dt – อัตราการผลิตเอนโทรปี V คือปริมาตรของระบบ
ฟังก์ชันการกระจายคือการวัดการกระจายพลังงานของระบบไปสู่ความร้อน จะกำหนดอัตราการเพิ่มขึ้นของเอนโทรปีในระบบซึ่งกระบวนการที่ไม่สามารถย้อนกลับได้เกิดขึ้น ยิ่งค่าของฟังก์ชันการกระจายสูงขึ้น พลังงานทุกประเภทจะถูกแปลงเป็นความร้อนเร็วขึ้น นอกจากนี้ ฟังก์ชันการกระจายจะกำหนดความเป็นไปได้ของกระบวนการที่เกิดขึ้นเอง: สำหรับ ψ>0 กระบวนการนั้นเป็นไปได้ สำหรับ ψ<0 – нет.
อุณหพลศาสตร์แสดงให้เห็นว่าหากระบบไม่มีความสมดุล แต่ใกล้กับสมดุล ดังนั้น ψ สามารถแทนได้ด้วยผลรวมของผลคูณของแรงทั่วไป - Xi และการไหลทั่วไป - Φi นั่นคือผลรวมของกำลังของกระบวนการ ψ = ∑ ΦiXi ≥0 ค่าบวกของฟังก์ชันการกระจาย ψ หมายความว่าในตัวแปลงพลังงานใดๆ กำลังไฟฟ้าเข้าจะต้องเกินกำลังไฟฟ้าเอาท์พุต ในกระบวนการทางชีวภาพส่วนใหญ่ พลังงานเคมีจะถูกแปลงเป็นพลังงานออสโมติก พลังงานไฟฟ้า และพลังงานกล ในกระบวนการทั้งหมดนี้ พลังงานเคมีส่วนหนึ่งจะกระจายไปเป็นความร้อน สำหรับกระบวนการทางชีววิทยา ประสิทธิภาพการเชื่อมต่อคือ 80-90% นั่นคือพลังงานเพียง 10-20% ที่เปลี่ยนเป็นความร้อน
สถานะคงที่ของระบบเปิดมีลักษณะเฉพาะตามทฤษฎีบทของ Prigogine: ในสถานะคงที่ซึ่งมีพารามิเตอร์ภายนอกคงที่ อัตราการผลิตเอนโทรปีในระบบจะคงที่ในเวลาและมีมูลค่าน้อยที่สุด
หากเกณฑ์สำหรับวิวัฒนาการของระบบในอุณหพลศาสตร์คลาสสิกคือ เอนโทรปีสำหรับกระบวนการที่ไม่สามารถย้อนกลับได้ในระบบที่แยกออกมามีแนวโน้มที่จะมีค่าสูงสุด ( เกณฑ์ของซานตาคลอส) จากนั้นในระบบเปิด การผลิตเอนโทรปีมีแนวโน้มน้อยที่สุด( เกณฑ์พรีโกจีน). เกณฑ์ Prigogine (Δψ>0) - เกณฑ์ความเสถียร - เมื่อเบี่ยงเบนไปจากสถานะเสถียร Δψ<0. Это является доказательством того, что второй закон термодинамики выполняется в живой природе.
จากทฤษฎีบทของ Prigogine จะได้ว่าหากระบบถูกนำออกจากสถานะหยุดนิ่ง ระบบก็จะเปลี่ยนแปลงไปจนกว่าอัตราการผลิตเอนโทรปีจำเพาะจะใช้ค่าที่ต่ำที่สุด นั่นคือจนกว่าฟังก์ชันการกระจายจะถึงค่าต่ำสุด
วิถีแห่งการแปลงพลังงานในเซลล์ที่มีชีวิตกลไกระดับโมเลกุลของปฏิกิริยาออกซิเดชันและฟอสโฟรีเลชั่นถูกถอดรหัสโดยมิตเชลล์ในปี 1976 ผู้เขียนได้พัฒนาทฤษฎีทางเคมีของออกซิเดชั่นฟอสโฟรีเลชั่น ส่วนที่สองของทฤษฎีของมิทเชลก็คือ มี ATPase ที่ไม่สมมาตรในเมมเบรนซึ่งทำงานแบบย้อนกลับได้ กล่าวคือ มันสามารถเป็น ATP synthetase ได้ด้วย:
ATP + NON (ATPase) ADP + P + 2H +
ความไม่สมดุลในการทำงานของ ATPase ก็คือ
ก) ในระหว่างการไฮโดรไลซิสของ ATP โปรตอน H+ และไฮดรอกซิล OH- จะถูกจับที่ด้านตรงข้ามของเมมเบรน
b) ในระหว่างการสังเคราะห์ ATP น้ำจะแยกตัวออกเป็น OH- ซึ่งเข้าสู่ด้านที่เป็นกรดของเมมเบรนมากกว่า และ H+ ซึ่งกระจายไปในทิศทางตรงกันข้าม
โดยทั่วไป กระบวนการของฟอสโฟรีเลชันของ ADP เกิดขึ้นเนื่องจากการเปลี่ยนแปลงของพลังงานอิสระระหว่างการทำให้ OH- ไอออนเป็นกลางในสภาพแวดล้อมที่เป็นกรด และไอออน H+ ในสภาพแวดล้อมที่เป็นด่าง
จากมุมมองของการแปลงพลังงานกระบวนการออกซิเดชั่นฟอสโฟรีเลชั่นประกอบด้วยสองขั้นตอน:
1. การแปลงพลังงานเคมีของการถ่ายโอนอิเล็กตรอนเป็นพลังงานที่เกี่ยวข้องกับความแตกต่างในศักย์ไฟฟ้าเคมีของโปรตอนอันเป็นผลมาจากการจับคู่ของการถ่ายโอนอิเล็กตรอนไปตามห่วงโซ่ทางเดินหายใจและการถ่ายโอนโปรตอนผ่านเมมเบรน ในกรณีนี้: Δμ H+ = FΔφ M + RT ln ( 1 / 2) โดยที่ Δμ H+ คือความแตกต่างในศักย์ไฟฟ้าเคมี Δφ M – ความแตกต่างของศักย์ไฟฟ้าระหว่างด้านนอกและด้านในของเยื่อหุ้มไมโตคอนเดรีย ( 1 และ 2 – ความเข้มข้นของโปรตอนในสิ่งแวดล้อมและภายในไมโตคอนเดรีย
2. การแปลงพลังงานที่กำหนดโดยความแตกต่างในศักย์ไฟฟ้าเป็นพลังงานเคมีของพันธะพลังงานสูงของ ATP (เชื่อมโยงการถ่ายโอน 2H+ และการสังเคราะห์ ATP หนึ่งโมเลกุลจาก ADP และฟอสเฟต) สิ่งนี้สามารถแสดงตามอัตภาพเป็น Δμ H+ → QUOTE ~ ~
ขณะนี้แสดงให้เห็นว่าในกรณีที่มีความแตกต่างในศักยภาพทางเคมีไฟฟ้า H+ ไม่เพียงแต่งานทางเคมี (การสังเคราะห์ ATP) แต่ยังรวมถึงงานออสโมติกด้วย (ระหว่างการขนส่งสารประกอบต่างๆ ผ่านเยื่อหุ้มเซลล์) งานทางกล (การเคลื่อนที่ของแฟลเจลลาในแบคทีเรีย) และความร้อนก็ถูกปล่อยออกมาเช่นกัน (การแยกส่วนการควบคุมความร้อนของออกซิเดชั่นฟอสโฟรีเลชั่น)
ในเชิงสัญลักษณ์ ทฤษฎีทางเคมีของการเชื่อมโยงกระบวนการออกซิเดชัน (เช่นการถ่ายโอนอิเล็กตรอน - e) และฟอสโฟรีเลชั่น (การสังเคราะห์ macroergs - QUOTE ~ ~) สามารถแสดงในรูปแบบของแผนภาพ e QUOTE Δμ H+ QUOTEอ้าง ~ ~. ผลลัพธ์หลักต่อไปนี้ของทฤษฎีเคมีบำบัดเป็นไปตามแผนนี้:
1. ถ้า Δμ H+ = 0 การสังเคราะห์ ATP จะไม่เกิดขึ้นระหว่างการถ่ายโอนอิเล็กตรอน
2. เมื่อระบบทางเดินหายใจทำงาน ศักยภาพของเมมเบรนจะถูกสร้างขึ้น (e→Δφ M)
3. การสร้างศักย์ไฟฟ้าที่เพียงพอบนเมมเบรนข้อต่อพลังงานที่มีเครื่องหมาย "+" ที่ด้านนอกจะนำไปสู่การสังเคราะห์ ATP จาก ADP และออร์โธฟอสเฟต (Δφ M → QUOTE ~) ~)
4. เนื่องจากศักยภาพของเมมเบรน จึงเป็นไปได้ที่จะหยุดและแม้แต่ "ย้อนกลับ" การไหลของอิเล็กตรอนในห่วงโซ่ทางเดินหายใจ (Δφ M →e)
5. เมื่อ ATP ถูกไฮโดรไลซ์ที่เมมเบรนข้อต่อ จะเกิดศักย์ไฟฟ้าของเมมเบรน (QUOTE ~ ~ → Δφ M)
ดังนั้นงานประเภทหลักในเซลล์ที่มีชีวิต - ไฟฟ้าและออสโมติก - จะดำเนินการโดยมีส่วนร่วมโดยตรงของเยื่อหุ้มชีวภาพ มีบทบาทสำคัญในพลังงานของเซลล์โดยกระบวนการสังเคราะห์และสลาย ATP ในเซลล์ ATP เป็นตัวสะสมพลังงานเคมี
พลังงานถูกใช้สำหรับปฏิกิริยาเคมีต่างๆ ที่เกิดขึ้นในเซลล์ สิ่งมีชีวิตบางชนิดใช้พลังงานของแสงแดดสำหรับกระบวนการทางชีวเคมี - สิ่งเหล่านี้คือพืช ในขณะที่สิ่งมีชีวิตบางชนิดใช้พลังงานของพันธะเคมีในสารที่ได้รับระหว่างโภชนาการ - สิ่งเหล่านี้คือสิ่งมีชีวิตของสัตว์ สารจากอาหารถูกสกัดโดยการสลายหรือออกซิเดชันทางชีวภาพโดยผ่านกระบวนการหายใจของเซลล์
การหายใจระดับเซลล์เป็นกระบวนการทางชีวเคมีในเซลล์ที่เกิดขึ้นเมื่อมีเอนไซม์ซึ่งเป็นผลมาจากการที่น้ำและคาร์บอนไดออกไซด์ถูกปล่อยออกมาพลังงานจะถูกเก็บไว้ในรูปของพันธะพลังงานมหภาคของโมเลกุล ATP หากกระบวนการนี้เกิดขึ้นโดยมีออกซิเจนอยู่ จะเรียกว่า "แอโรบิก" ถ้ามันเกิดขึ้นโดยไม่มีออกซิเจน เรียกว่า “แอนแอโรบิก”
ออกซิเดชันทางชีวภาพประกอบด้วยสามขั้นตอนหลัก:
1. เตรียมความพร้อม
2. ปราศจากออกซิเจน (ไกลโคไลซิส)
3. การสลายสารอินทรีย์โดยสมบูรณ์ (เมื่อมีออกซิเจน)
ขั้นตอนการเตรียมการ สารที่ได้รับจากอาหารจะถูกแบ่งออกเป็นโมโนเมอร์ ระยะนี้เริ่มต้นในทางเดินอาหารหรือในไลโซโซมของเซลล์ โพลีแซ็กคาไรด์แตกตัวเป็นโมโนแซ็กคาไรด์ โปรตีนเป็นกรดอะมิโน ไขมันเป็นกลีเซอรอลและกรดไขมัน พลังงานที่ปล่อยออกมาในขั้นตอนนี้จะกระจายไปในรูปของความร้อน ควรสังเกตว่าสำหรับกระบวนการพลังงาน เซลล์ใช้คาร์โบไฮเดรต หรือดีกว่านั้นคือโมโนแซ็กคาไรด์ และสมองสามารถใช้เพียงโมโนแซ็กคาไรด์ - กลูโคส - ในการทำงานเท่านั้น
กลูโคสในระหว่างไกลโคไลซิสจะแบ่งออกเป็นโมเลกุลสามคาร์บอนของกรดไพรูวิค ชะตากรรมต่อไปของพวกเขาขึ้นอยู่กับการมีอยู่ของออกซิเจนในเซลล์ หากมีออกซิเจนอยู่ในเซลล์ กรดไพรูวิกจะเข้าสู่ไมโตคอนเดรียเพื่อเกิดออกซิเดชันอย่างสมบูรณ์กับคาร์บอนไดออกไซด์และน้ำ (การหายใจแบบใช้ออกซิเจน) หากไม่มีออกซิเจน กรดไพรูวิกในเนื้อเยื่อของสัตว์จะถูกเปลี่ยนเป็นกรดแลคติค ระยะนี้เกิดขึ้นในไซโตพลาสซึมของเซลล์ จากผลของไกลโคไลซิส จะเกิดโมเลกุล ATP เพียง 2 โมเลกุลเท่านั้น
หากต้องการออกซิเดชันกลูโคสโดยสมบูรณ์ จำเป็นต้องมีออกซิเจน ในระยะที่สาม ปฏิกิริยาออกซิเดชันที่สมบูรณ์ของกรดไพรูวิกเป็นคาร์บอนไดออกไซด์และน้ำเกิดขึ้นในไมโตคอนเดรีย เป็นผลให้เกิดโมเลกุล ATP อีก 36 โมเลกุล
โดยรวมแล้ว ขั้นตอนทั้งสามนี้สร้างโมเลกุล ATP ได้ 38 ATP จากโมเลกุลกลูโคส 1 โมเลกุล โดยคำนึงถึง ATP 2 ตัวที่เกิดขึ้นระหว่างไกลโคไลซิส
ดังนั้นเราจึงตรวจสอบกระบวนการพลังงานที่เกิดขึ้นในเซลล์ ระยะของการเกิดออกซิเดชันทางชีวภาพมีลักษณะเฉพาะ นี่เป็นการสรุปบทเรียนของเรา ขอให้โชคดี ลาก่อน!
ความแตกต่างระหว่างการหายใจและการเผาไหม้. การหายใจที่เกิดขึ้นในเซลล์มักถูกเปรียบเทียบกับกระบวนการเผาไหม้ กระบวนการทั้งสองเกิดขึ้นในที่ที่มีออกซิเจน โดยปล่อยพลังงานและผลิตภัณฑ์ออกซิเดชัน แต่แตกต่างจากการเผาไหม้ตรงที่การหายใจเป็นกระบวนการที่ได้รับคำสั่งของปฏิกิริยาทางชีวเคมีที่เกิดขึ้นเมื่อมีเอนไซม์ ในระหว่างการหายใจ คาร์บอนไดออกไซด์จะเกิดขึ้นเป็นผลสุดท้ายของการเกิดออกซิเดชันทางชีวภาพ และในระหว่างการเผาไหม้ การก่อตัวของก๊าซคาร์บอนไดออกไซด์เกิดขึ้นจากการรวมตัวโดยตรงของไฮโดรเจนกับคาร์บอน นอกจากนี้ในระหว่างการหายใจจะเกิดโมเลกุล ATP จำนวนหนึ่งเกิดขึ้น นั่นคือการหายใจและการเผาไหม้เป็นกระบวนการที่แตกต่างกันโดยพื้นฐาน
ความสำคัญทางชีวการแพทย์สำหรับการแพทย์ ไม่เพียงแต่การเผาผลาญกลูโคสเท่านั้นที่สำคัญ แต่ยังรวมถึงฟรุกโตสและกาแลคโตสด้วย ความสามารถในการสร้าง ATP ในกรณีที่ไม่มีออกซิเจนมีความสำคัญอย่างยิ่งในทางการแพทย์ สิ่งนี้ช่วยให้คุณรักษาการทำงานของกล้ามเนื้อโครงร่างได้อย่างเข้มข้นในสภาวะที่ประสิทธิภาพการออกซิเดชั่นแบบแอโรบิกไม่เพียงพอ เนื้อเยื่อที่มีกิจกรรมไกลโคไลติกเพิ่มขึ้นสามารถยังคงทำงานต่อไปได้ในช่วงที่ขาดออกซิเจน ในกล้ามเนื้อหัวใจ ความเป็นไปได้ของไกลโคไลซิสมีจำกัด เธอมีช่วงเวลาที่ยากลำบากจากการหยุดชะงักของการจัดหาเลือด ซึ่งอาจนำไปสู่ภาวะขาดเลือดได้ มีโรคหลายชนิดที่เกิดจากการขาดเอนไซม์ที่ควบคุมไกลโคไลซิส:
- โรคโลหิตจางจากเม็ดเลือดแดงแตก (ในเซลล์มะเร็งที่เติบโตอย่างรวดเร็ว glycolysis เกิดขึ้นในอัตราที่เกินความสามารถของวงจรกรดซิตริก) ซึ่งมีส่วนช่วยในการสังเคราะห์กรดแลคติคในอวัยวะและเนื้อเยื่อเพิ่มขึ้น กรดแลคติคในร่างกายในปริมาณที่สูงอาจเป็นสัญญาณของมะเร็งได้
การหมักจุลินทรีย์สามารถรับพลังงานได้ในระหว่างการหมัก ผู้คนรู้จักการหมักมาตั้งแต่สมัยโบราณ เช่น ในการผลิตไวน์ การหมักกรดแลกติกเป็นที่รู้จักก่อนหน้านี้ด้วยซ้ำ ผู้คนบริโภคผลิตภัณฑ์นมโดยไม่รู้ว่ากระบวนการเหล่านี้เกี่ยวข้องกับกิจกรรมของจุลินทรีย์ สิ่งนี้ได้รับการพิสูจน์ครั้งแรกโดยหลุยส์ ปาสเตอร์ ยิ่งไปกว่านั้น จุลินทรีย์ต่าง ๆ ยังหลั่งผลิตภัณฑ์จากการหมักที่แตกต่างกันออกไป ตอนนี้เราจะพูดถึงการหมักแอลกอฮอล์และกรดแลคติค เป็นผลให้เอทิลแอลกอฮอล์และคาร์บอนไดออกไซด์เกิดขึ้นและปล่อยพลังงานออกมา ผู้ผลิตเบียร์และผู้ผลิตไวน์ใช้ยีสต์บางประเภทเพื่อกระตุ้นการหมัก ซึ่งจะเปลี่ยนน้ำตาลให้เป็นแอลกอฮอล์ การหมักส่วนใหญ่ดำเนินการโดยยีสต์ เช่นเดียวกับแบคทีเรียและเชื้อราบางชนิด ในประเทศของเรา มีการใช้ยีสต์ Saccharomycetes แบบดั้งเดิม ในอเมริกา - แบคทีเรียในสกุล Pseudomonas และในเม็กซิโก มีการใช้แบคทีเรีย "ก้านเคลื่อนที่" โดยทั่วไปยีสต์ของเราหมักเฮกโซส (โมโนแซ็กคาไรด์หกคาร์บอน) เช่น กลูโคสหรือฟรุกโตส กระบวนการสร้างแอลกอฮอล์สามารถแสดงได้ดังนี้: จากโมเลกุลกลูโคสหนึ่งโมเลกุลแอลกอฮอล์สองโมเลกุลคาร์บอนไดออกไซด์สองโมเลกุลและ ATP สองโมเลกุลเกิดขึ้น วิธีนี้มีกำไรน้อยกว่ากระบวนการแอโรบิก แต่ช่วยให้คุณรักษาชีวิตไว้ได้หากไม่มีออกซิเจน ทีนี้มาพูดถึงการหมักนมเปรี้ยวกันดีกว่า กลูโคสหนึ่งโมเลกุลก่อให้เกิดกรดแลคติคสองโมเลกุลและในเวลาเดียวกันก็ปล่อย ATP สองโมเลกุลออกมา การหมักกรดแลกติกถูกนำมาใช้กันอย่างแพร่หลายในการผลิตผลิตภัณฑ์นม: ชีส นมเปรี้ยว โยเกิร์ต กรดแลคติคยังใช้ในการผลิตน้ำอัดลม
ความสามารถในการสังเคราะห์แสงเป็นลักษณะสำคัญของพืชสีเขียว พืชก็เหมือนกับสิ่งมีชีวิตทุกชนิดที่ต้องมี กิน หายใจ กำจัดสารที่ไม่จำเป็น เติบโต สืบพันธุ์ ตอบสนองต่อการเปลี่ยนแปลงของสิ่งแวดล้อม. ทั้งหมดนี้รับประกันได้จากการทำงานของอวัยวะที่เกี่ยวข้องของร่างกาย โดยปกติแล้ว อวัยวะต่างๆ จะสร้างระบบของอวัยวะที่ทำงานร่วมกันเพื่อให้มั่นใจในการทำงานของสิ่งมีชีวิตอย่างใดอย่างหนึ่งหรืออย่างอื่น ดังนั้นสิ่งมีชีวิตจึงสามารถแสดงเป็นระบบชีวภาพได้ แต่ละอวัยวะในพืชที่มีชีวิตทำหน้าที่เฉพาะเจาะจง รากดูดซับน้ำด้วยแร่ธาตุจากดินและทำให้พืชในดินแข็งแรง ก้านใบจะพาใบไม้เข้าหาแสง น้ำตลอดจนแร่ธาตุและสารอินทรีย์เคลื่อนที่ไปตามก้าน ในคลอโรพลาสต์ของใบไม้ภายใต้แสง สารอินทรีย์จะเกิดขึ้นจากสารอนินทรีย์ที่พวกมันกินเข้าไป เซลล์อวัยวะทั้งหมด พืช. ใบไม้ระเหยน้ำ
หากการทำงานของอวัยวะส่วนใดส่วนหนึ่งในร่างกายถูกรบกวน อาจส่งผลให้การทำงานของอวัยวะอื่นๆ และทั้งร่างกายหยุดชะงักได้ ตัวอย่างเช่น หากน้ำหยุดไหลผ่านราก ต้นไม้ทั้งต้นก็อาจตายได้ หากพืชผลิตคลอโรฟิลล์ในใบไม่เพียงพอ ก็จะไม่สามารถสังเคราะห์สารอินทรีย์ในปริมาณที่เพียงพอต่อการทำงานที่สำคัญของมันได้
ดังนั้นกิจกรรมที่สำคัญของร่างกายจึงมั่นใจได้จากการทำงานที่เชื่อมโยงถึงกันของทุกระบบอวัยวะ กิจกรรมในชีวิตคือกระบวนการทั้งหมดที่เกิดขึ้นในร่างกาย
ต้องขอบคุณโภชนาการที่ทำให้ร่างกายมีชีวิตและเติบโต ในระหว่างโภชนาการ สารที่จำเป็นจะถูกดูดซึมจากสิ่งแวดล้อม จากนั้นจะถูกดูดซึมเข้าสู่ร่างกาย พืชดูดซับน้ำและแร่ธาตุจากดิน อวัยวะสีเขียวเหนือพื้นดินของพืชดูดซับคาร์บอนไดออกไซด์จากอากาศ พืชใช้น้ำและคาร์บอนไดออกไซด์เพื่อสังเคราะห์สารอินทรีย์ ซึ่งพืชใช้ในการสร้างเซลล์ใหม่ เจริญเติบโต และพัฒนา
การแลกเปลี่ยนก๊าซเกิดขึ้นระหว่างการหายใจ ออกซิเจนถูกดูดซับจากสิ่งแวดล้อม และก๊าซคาร์บอนไดออกไซด์และไอน้ำถูกปล่อยออกมาจากร่างกาย เซลล์ที่มีชีวิตทั้งหมดต้องการออกซิเจนเพื่อผลิตพลังงาน
ในระหว่างกระบวนการเผาผลาญ สารที่ร่างกายไม่ต้องการจะถูกสร้างและปล่อยออกสู่สิ่งแวดล้อม
เมื่อพืชมีอายุถึงขนาดและอายุที่จำเป็นสำหรับสายพันธุ์ของมัน หากอยู่ในสภาพแวดล้อมที่เอื้ออำนวยเพียงพอ มันก็จะเริ่มแพร่พันธุ์ อันเป็นผลมาจากการสืบพันธุ์ทำให้จำนวนบุคคลเพิ่มขึ้น
พืชต่างจากสัตว์ส่วนใหญ่ที่เติบโตตลอดชีวิต
การได้มาซึ่งคุณสมบัติใหม่จากสิ่งมีชีวิตเรียกว่าการพัฒนา
โภชนาการ การหายใจ เมแทบอลิซึม การเจริญเติบโตและพัฒนาการ รวมถึงการสืบพันธุ์ได้รับอิทธิพลจากสภาพแวดล้อมของพืช หากพวกมันไม่ดีพอ พืชก็อาจเติบโตและพัฒนาได้ไม่ดี กระบวนการสำคัญของมันจะถูกระงับ ดังนั้นชีวิตของพืชจึงขึ้นอยู่กับสภาพแวดล้อม
คำถามที่ 3_เยื่อหุ้มเซลล์ หน้าที่ องค์ประกอบ โครงสร้าง เปลือกหลักและรอง
เซลล์ของสิ่งมีชีวิตใดๆ ก็ตามเป็นระบบที่มีชีวิตครบถ้วน ประกอบด้วยสามส่วนที่เชื่อมโยงกันอย่างแยกไม่ออก: เมมเบรน, ไซโตพลาสซึมและนิวเคลียส เยื่อหุ้มเซลล์โต้ตอบโดยตรงกับสภาพแวดล้อมภายนอกและโต้ตอบกับเซลล์ข้างเคียง (ในสิ่งมีชีวิตหลายเซลล์) เยื่อหุ้มเซลล์. เยื่อหุ้มเซลล์มีโครงสร้างที่ซับซ้อน ประกอบด้วยชั้นนอกและพลาสมาเมมเบรนที่อยู่ข้างใต้ ในพืช เช่นเดียวกับในแบคทีเรีย สาหร่ายและเชื้อราสีน้ำเงินแกมเขียว เมมเบรนหนาแน่น หรือผนังเซลล์จะตั้งอยู่บนพื้นผิวของเซลล์ ในพืชส่วนใหญ่ประกอบด้วยเส้นใย ผนังเซลล์มีบทบาทสำคัญอย่างยิ่ง โดยเป็นโครงด้านนอก เป็นเกราะป้องกัน และทำหน้าที่เป็นเทอร์กอร์ให้กับเซลล์พืช น้ำ เกลือ และโมเลกุลของสารอินทรีย์หลายชนิดทะลุผ่านผนังเซลล์
เยื่อหุ้มเซลล์หรือผนัง - เยื่อหุ้มเซลล์แข็งซึ่งอยู่นอกเยื่อหุ้มเซลล์ไซโตพลาสซึมและทำหน้าที่ด้านโครงสร้าง การป้องกัน และการขนส่ง พบได้ในแบคทีเรีย อาร์เคีย เชื้อรา และพืชส่วนใหญ่ สัตว์และโปรโตซัวหลายชนิดไม่มีผนังเซลล์
หน้าที่ของเยื่อหุ้มเซลล์:
1. ฟังก์ชั่นการขนส่งให้การควบคุมแบบเลือกสรรของการเผาผลาญระหว่างเซลล์และสภาพแวดล้อมภายนอก การไหลของสารเข้าสู่เซลล์ (เนื่องจากการซึมผ่านของเยื่อหุ้มเซลล์แบบกึ่งซึมผ่าน) รวมถึงการควบคุมสมดุลของน้ำของเซลล์
1.1. การเคลื่อนย้ายเมมเบรน (เช่น ข้ามเมมเบรน):
- การแพร่กระจาย
- การขนส่งแบบพาสซีฟ = การอำนวยความสะดวกในการแพร่กระจาย
- Active = การขนส่งแบบเลือกสรร (เกี่ยวข้องกับ ATP และเอนไซม์)
1.2. การขนส่งในบรรจุภัณฑ์เมมเบรน:
- Exocytosis - การปล่อยสารออกจากเซลล์
- Endocytosis (phago- และ pinocytosis) - การดูดซึมสารโดยเซลล์
2) ฟังก์ชั่นตัวรับ.
3) การสนับสนุน (“โครงกระดูก”)- คงรูปร่างของเซลล์ให้แข็งแรง นี่เป็นหน้าที่หลักของผนังเซลล์
4) การแยกเซลล์(สิ่งที่มีชีวิต) จากสิ่งแวดล้อม
5) ฟังก์ชั่นการป้องกัน
6) ติดต่อกับเซลล์ข้างเคียง การรวมกันของเซลล์เป็นเนื้อเยื่อ.
บทเรียนวิดีโอนี้เน้นในหัวข้อ “การให้พลังงานแก่เซลล์” ในบทนี้ เราจะดูกระบวนการพลังงานในเซลล์และศึกษาว่าเซลล์ได้รับพลังงานอย่างไร นอกจากนี้คุณยังจะได้เรียนรู้ว่าการหายใจของเซลล์คืออะไรและประกอบด้วยขั้นตอนใด พูดคุยรายละเอียดแต่ละขั้นตอนเหล่านี้
ชีววิทยา ชั้นประถมศึกษาปีที่ 9
หัวข้อ: ระดับเซลล์
บทที่ 13. ให้พลังงานแก่เซลล์
สเตปาโนวา แอนนา ยูริเยฟนา
ผู้สมัครสาขาวิทยาศาสตร์ชีวภาพรองศาสตราจารย์ มกุย
มอสโก
วันนี้เราจะมาพูดถึงการให้พลังงานแก่เซลล์ พลังงานถูกใช้สำหรับปฏิกิริยาเคมีต่างๆ ที่เกิดขึ้นในเซลล์ สิ่งมีชีวิตบางชนิดใช้พลังงานของแสงแดดสำหรับกระบวนการทางชีวเคมี - สิ่งเหล่านี้คือพืช ในขณะที่สิ่งมีชีวิตบางชนิดใช้พลังงานของพันธะเคมีในสารที่ได้รับระหว่างโภชนาการ - สิ่งเหล่านี้คือสิ่งมีชีวิตของสัตว์ สารจากอาหารถูกสกัดโดยการสลายหรือออกซิเดชันทางชีวภาพโดยผ่านกระบวนการหายใจของเซลล์
การหายใจระดับเซลล์เป็นกระบวนการทางชีวเคมีในเซลล์ที่เกิดขึ้นเมื่อมีเอนไซม์ซึ่งเป็นผลมาจากการที่น้ำและคาร์บอนไดออกไซด์ถูกปล่อยออกมาพลังงานจะถูกเก็บไว้ในรูปของพันธะพลังงานมหภาคของโมเลกุล ATP หากกระบวนการนี้เกิดขึ้นโดยมีออกซิเจนอยู่ จะเรียกว่า "แอโรบิก" ถ้ามันเกิดขึ้นโดยไม่มีออกซิเจน เรียกว่า “แอนแอโรบิก”
ออกซิเดชันทางชีวภาพประกอบด้วยสามขั้นตอนหลัก:
1. เตรียมการ
2. ปราศจากออกซิเจน (ไกลโคไลซิส)
3. การสลายสารอินทรีย์โดยสมบูรณ์ (เมื่อมีออกซิเจน)
ขั้นตอนการเตรียมการ สารที่ได้รับจากอาหารจะถูกแบ่งออกเป็นโมโนเมอร์ ระยะนี้เริ่มต้นในทางเดินอาหารหรือในไลโซโซมของเซลล์ โพลีแซ็กคาไรด์แตกตัวเป็นโมโนแซ็กคาไรด์ โปรตีนเป็นกรดอะมิโน ไขมันเป็นกลีเซอรอลและกรดไขมัน พลังงานที่ปล่อยออกมาในขั้นตอนนี้จะกระจายไปในรูปของความร้อน ควรสังเกตว่าสำหรับกระบวนการพลังงาน เซลล์ใช้คาร์โบไฮเดรต หรือดีกว่านั้นคือโมโนแซ็กคาไรด์ และสมองสามารถใช้เพียงโมโนแซ็กคาไรด์ - กลูโคส - ในการทำงานเท่านั้น
กลูโคสในระหว่างไกลโคไลซิสจะแบ่งออกเป็นโมเลกุลสามคาร์บอนของกรดไพรูวิค ชะตากรรมต่อไปของพวกเขาขึ้นอยู่กับการมีอยู่ของออกซิเจนในเซลล์ หากมีออกซิเจนอยู่ในเซลล์ กรดไพรูวิกจะเข้าสู่ไมโตคอนเดรียเพื่อเกิดออกซิเดชันอย่างสมบูรณ์กับคาร์บอนไดออกไซด์และน้ำ (การหายใจแบบใช้ออกซิเจน) หากไม่มีออกซิเจน กรดไพรูวิกในเนื้อเยื่อของสัตว์จะถูกเปลี่ยนเป็นกรดแลคติค ระยะนี้เกิดขึ้นในไซโตพลาสซึมของเซลล์ จากผลของไกลโคไลซิส จะเกิดโมเลกุล ATP เพียง 2 โมเลกุลเท่านั้น
หากต้องการออกซิเดชันกลูโคสโดยสมบูรณ์ จำเป็นต้องมีออกซิเจน ในระยะที่สาม ปฏิกิริยาออกซิเดชันที่สมบูรณ์ของกรดไพรูวิกเป็นคาร์บอนไดออกไซด์และน้ำเกิดขึ้นในไมโตคอนเดรีย เป็นผลให้เกิดโมเลกุล ATP อีก 36 โมเลกุล
โดยรวมแล้ว ขั้นตอนทั้งสามนี้สร้างโมเลกุล ATP ได้ 38 ATP จากโมเลกุลกลูโคส 1 โมเลกุล โดยคำนึงถึง ATP 2 ตัวที่เกิดขึ้นระหว่างไกลโคไลซิส
ดังนั้นเราจึงตรวจสอบกระบวนการพลังงานที่เกิดขึ้นในเซลล์ ระยะของการเกิดออกซิเดชันทางชีวภาพมีลักษณะเฉพาะ นี่เป็นการสรุปบทเรียนของเรา ขอให้โชคดี ลาก่อน!
ความแตกต่างระหว่างการหายใจและการเผาไหม้. การหายใจที่เกิดขึ้นในเซลล์มักถูกเปรียบเทียบกับกระบวนการเผาไหม้ กระบวนการทั้งสองเกิดขึ้นในที่ที่มีออกซิเจน โดยปล่อยพลังงานและผลิตภัณฑ์ออกซิเดชัน แต่แตกต่างจากการเผาไหม้ตรงที่การหายใจเป็นกระบวนการที่ได้รับคำสั่งของปฏิกิริยาทางชีวเคมีที่เกิดขึ้นเมื่อมีเอนไซม์ ในระหว่างการหายใจ คาร์บอนไดออกไซด์จะเกิดขึ้นเป็นผลสุดท้ายของการเกิดออกซิเดชันทางชีวภาพ และในระหว่างการเผาไหม้ การก่อตัวของก๊าซคาร์บอนไดออกไซด์เกิดขึ้นจากการรวมตัวโดยตรงของไฮโดรเจนกับคาร์บอน นอกจากนี้ในระหว่างการหายใจจะเกิดโมเลกุล ATP จำนวนหนึ่งเกิดขึ้น นั่นคือการหายใจและการเผาไหม้เป็นกระบวนการที่แตกต่างกันโดยพื้นฐาน
ความสำคัญทางชีวการแพทย์สำหรับการแพทย์ ไม่เพียงแต่การเผาผลาญกลูโคสเท่านั้นที่สำคัญ แต่ยังรวมถึงฟรุกโตสและกาแลคโตสด้วย ความสามารถในการสร้าง ATP ในกรณีที่ไม่มีออกซิเจนมีความสำคัญอย่างยิ่งในทางการแพทย์ สิ่งนี้ช่วยให้คุณรักษาการทำงานของกล้ามเนื้อโครงร่างได้อย่างเข้มข้นในสภาวะที่ประสิทธิภาพการออกซิเดชั่นแบบแอโรบิกไม่เพียงพอ เนื้อเยื่อที่มีกิจกรรมไกลโคไลติกเพิ่มขึ้นสามารถยังคงทำงานต่อไปได้ในช่วงที่ขาดออกซิเจน ในกล้ามเนื้อหัวใจ ความเป็นไปได้ของไกลโคไลซิสมีจำกัด เธอมีช่วงเวลาที่ยากลำบากจากการหยุดชะงักของการจัดหาเลือด ซึ่งอาจนำไปสู่ภาวะขาดเลือดได้ มีโรคหลายชนิดที่เกิดจากการขาดเอนไซม์ที่ควบคุมไกลโคไลซิส:
โรคโลหิตจางจากเม็ดเลือดแดงแตก (ในเซลล์มะเร็งที่เติบโตอย่างรวดเร็ว glycolysis เกิดขึ้นในอัตราที่เกินความสามารถของวงจรกรดซิตริก) ซึ่งมีส่วนช่วยในการสังเคราะห์กรดแลคติคในอวัยวะและเนื้อเยื่อเพิ่มขึ้น กรดแลคติคในร่างกายในปริมาณที่สูงอาจเป็นสัญญาณของมะเร็งได้
การหมักจุลินทรีย์สามารถรับพลังงานได้ในระหว่างการหมัก ผู้คนรู้จักการหมักมาตั้งแต่สมัยโบราณ เช่น ในการผลิตไวน์ การหมักกรดแลกติกเป็นที่รู้จักก่อนหน้านี้ด้วยซ้ำ ผู้คนบริโภคผลิตภัณฑ์นมโดยไม่รู้ว่ากระบวนการเหล่านี้เกี่ยวข้องกับกิจกรรมของจุลินทรีย์ สิ่งนี้ได้รับการพิสูจน์ครั้งแรกโดยหลุยส์ ปาสเตอร์ ยิ่งไปกว่านั้น จุลินทรีย์ต่าง ๆ ยังหลั่งผลิตภัณฑ์จากการหมักที่แตกต่างกันออกไป ตอนนี้เราจะพูดถึงการหมักแอลกอฮอล์และกรดแลคติค เป็นผลให้เอทิลแอลกอฮอล์และคาร์บอนไดออกไซด์เกิดขึ้นและปล่อยพลังงานออกมา ผู้ผลิตเบียร์และผู้ผลิตไวน์ใช้ยีสต์บางประเภทเพื่อกระตุ้นการหมัก ซึ่งจะเปลี่ยนน้ำตาลให้เป็นแอลกอฮอล์ การหมักส่วนใหญ่ดำเนินการโดยยีสต์ เช่นเดียวกับแบคทีเรียและเชื้อราบางชนิด ในประเทศของเรา มีการใช้ยีสต์ Saccharomycetes แบบดั้งเดิม ในอเมริกา - แบคทีเรียในสกุล Pseudomonas และในเม็กซิโก มีการใช้แบคทีเรีย "ก้านเคลื่อนที่" โดยทั่วไปยีสต์ของเราหมักเฮกโซส (โมโนแซ็กคาไรด์หกคาร์บอน) เช่น กลูโคสหรือฟรุกโตส กระบวนการสร้างแอลกอฮอล์สามารถแสดงได้ดังนี้: จากโมเลกุลกลูโคสหนึ่งโมเลกุลแอลกอฮอล์สองโมเลกุลคาร์บอนไดออกไซด์สองโมเลกุลและ ATP สองโมเลกุลเกิดขึ้น วิธีนี้มีกำไรน้อยกว่ากระบวนการแอโรบิก แต่ช่วยให้คุณรักษาชีวิตไว้ได้หากไม่มีออกซิเจน ทีนี้มาพูดถึงการหมักนมเปรี้ยวกันดีกว่า กลูโคสหนึ่งโมเลกุลก่อให้เกิดกรดแลคติคสองโมเลกุลและในเวลาเดียวกันก็ปล่อย ATP สองโมเลกุลออกมา การหมักกรดแลกติกถูกนำมาใช้กันอย่างแพร่หลายในการผลิตผลิตภัณฑ์นม: ชีส นมเปรี้ยว โยเกิร์ต กรดแลคติคยังใช้ในการผลิตน้ำอัดลม
การเจริญเติบโตของไม้อ้วนมากมาย
ซึ่งหยั่งรากอยู่บนผืนทรายอันแห้งแล้ง
อนุมัติระบุชัดเจนว่า
แผ่นไขมัน ไขมันจากอากาศ
ดูดซับ...
เอ็ม.วี. โลโมโนซอฟ
พลังงานถูกเก็บไว้ในเซลล์อย่างไร? เมแทบอลิซึมคืออะไร? สาระสำคัญของกระบวนการไกลโคไลซิส การหมัก และการหายใจของเซลล์คืออะไร? กระบวนการใดเกิดขึ้นระหว่างช่วงแสงและช่วงมืดของการสังเคราะห์ด้วยแสง? กระบวนการเมแทบอลิซึมของพลังงานและพลาสติกเกี่ยวข้องกันอย่างไร? การสังเคราะห์ทางเคมีคืออะไร?
บทเรียนบรรยาย
ความสามารถในการแปลงพลังงานประเภทหนึ่งไปเป็นพลังงานอีกประเภทหนึ่ง (พลังงานรังสีเป็นพลังงานของพันธะเคมี พลังงานเคมีเป็นพลังงานกล ฯลฯ) เป็นหนึ่งในคุณสมบัติพื้นฐานของสิ่งมีชีวิต ที่นี่เราจะมาดูอย่างใกล้ชิดว่ากระบวนการเหล่านี้เกิดขึ้นได้อย่างไรในสิ่งมีชีวิต
ATP เป็นผู้ขนส่งพลังงานหลักภายในเซลล์. ในการดำเนินกิจกรรมของเซลล์จำเป็นต้องมีพลังงาน สิ่งมีชีวิตออโตโทรฟิคได้รับพลังงานเริ่มต้นจากดวงอาทิตย์ในระหว่างปฏิกิริยาสังเคราะห์ด้วยแสง ในขณะที่สิ่งมีชีวิตเฮเทอโรโทรฟิคใช้สารประกอบอินทรีย์ที่มาพร้อมกับอาหารเป็นแหล่งพลังงาน พลังงานถูกเก็บไว้ในเซลล์ในพันธะเคมีของโมเลกุล เอทีพี (อะดีโนซีน ไตรฟอสเฟต)ซึ่งเป็นนิวคลีโอไทด์ที่ประกอบด้วยกลุ่มฟอสเฟต 3 กลุ่ม กากน้ำตาล (ไรโบส) และกากเบสไนโตรเจน (อะดีนีน) (รูปที่ 52)
ข้าว. 52. โมเลกุลเอทีพี
พันธะระหว่างฟอสเฟตที่ตกค้างเรียกว่า Macroergic เนื่องจากเมื่อแตกตัวพลังงานจำนวนมากจะถูกปล่อยออกมา โดยปกติแล้ว เซลล์จะดึงพลังงานจาก ATP โดยการกำจัดเฉพาะกลุ่มฟอสเฟตส่วนปลายเท่านั้น ในกรณีนี้ จะเกิด ADP (อะดีโนซีน ไดฟอสเฟต) และกรดฟอสฟอริก และ 40 กิโลจูล/โมลถูกปล่อยออกมา:
โมเลกุลเอทีพีมีบทบาทเป็นชิปต่อรองพลังงานสากลของเซลล์ พวกเขาจะถูกส่งไปยังที่ตั้งของกระบวนการที่ใช้พลังงานมากไม่ว่าจะเป็นการสังเคราะห์เอนไซม์ของสารประกอบอินทรีย์การทำงานของโปรตีน - มอเตอร์โมเลกุลหรือโปรตีนการขนส่งเมมเบรน ฯลฯ การสังเคราะห์แบบย้อนกลับของโมเลกุล ATP นั้นดำเนินการโดยการแนบกลุ่มฟอสเฟต ไปจนถึง ADP ด้วยการดูดซับพลังงาน เซลล์เก็บพลังงานในรูปของ ATP ระหว่างปฏิกิริยา การเผาผลาญพลังงาน. มันมีความเกี่ยวข้องอย่างใกล้ชิดกับ การแลกเปลี่ยนพลาสติกในระหว่างที่เซลล์ผลิตสารประกอบอินทรีย์ที่จำเป็นสำหรับการทำงานของมัน
เมแทบอลิซึมและพลังงานในเซลล์ (เมแทบอลิซึม). เมแทบอลิซึมคือผลรวมของปฏิกิริยาทั้งหมดของเมแทบอลิซึมของพลาสติกและพลังงานที่เชื่อมโยงถึงกัน เซลล์สังเคราะห์คาร์โบไฮเดรต ไขมัน โปรตีน และกรดนิวคลีอิกอย่างต่อเนื่อง การสังเคราะห์สารประกอบมักเกิดขึ้นกับการใช้พลังงานที่สิ้นเปลืองเช่น ด้วยการมีส่วนร่วมที่ขาดไม่ได้ของ ATP แหล่งพลังงานสำหรับการก่อตัวของ ATP คือปฏิกิริยาของเอนไซม์ของการเกิดออกซิเดชันของโปรตีน ไขมัน และคาร์โบไฮเดรตที่เข้าสู่เซลล์ ในระหว่างกระบวนการนี้ พลังงานจะถูกปล่อยออกมาและเก็บไว้ใน ATP ออกซิเดชันของกลูโคสมีบทบาทพิเศษในการเผาผลาญพลังงานของเซลล์ โมเลกุลของกลูโคสได้รับการเปลี่ยนแปลงต่อเนื่องกันหลายครั้ง
ระยะแรกเรียกว่า ไกลโคไลซิสเกิดขึ้นในไซโตพลาสซึมของเซลล์และไม่ต้องใช้ออกซิเจน จากปฏิกิริยาต่อเนื่องที่เกี่ยวข้องกับเอนไซม์ กลูโคสจึงแตกตัวออกเป็นสองโมเลกุลของกรดไพรูวิก ในกรณีนี้ โมเลกุล ATP สองตัวถูกใช้ไป และพลังงานที่ปล่อยออกมาระหว่างการเกิดออกซิเดชันก็เพียงพอแล้วที่จะสร้างโมเลกุล ATP สี่โมเลกุล เป็นผลให้พลังงานที่ส่งออกของไกลโคไลซิสมีขนาดเล็กและมีจำนวนโมเลกุล ATP สองโมเลกุล:
ค 6 H1 2 0 6 → 2C 3 H 4 0 3 + 4H + + 2ATP
ภายใต้สภาวะไร้ออกซิเจน (ในกรณีที่ไม่มีออกซิเจน) การเปลี่ยนแปลงเพิ่มเติมสามารถเชื่อมโยงกับประเภทต่างๆ ได้ การหมัก.
ทุกคนรู้ การหมักกรดแลคติค(นมเปรี้ยว) ซึ่งเกิดขึ้นเนื่องจากการทำงานของเชื้อราและแบคทีเรียกรดแลคติค กลไกนี้คล้ายกับไกลโคไลซิส เพียงแต่ผลิตภัณฑ์สุดท้ายคือกรดแลคติค ปฏิกิริยาออกซิเดชันของกลูโคสประเภทนี้เกิดขึ้นในเซลล์เมื่อขาดออกซิเจน เช่น ในกล้ามเนื้อที่ทำงานอย่างเข้มข้น การหมักแอลกอฮอล์มีความใกล้เคียงกับการหมักกรดแลคติคในทางเคมี ความแตกต่างก็คือผลิตภัณฑ์จากการหมักแอลกอฮอล์คือเอทิลแอลกอฮอล์และคาร์บอนไดออกไซด์
ขั้นต่อไปในระหว่างที่กรดไพรูวิกถูกออกซิไดซ์เป็นคาร์บอนไดออกไซด์และน้ำเรียกว่า การหายใจของเซลล์. ปฏิกิริยาที่เกี่ยวข้องกับการหายใจเกิดขึ้นในไมโตคอนเดรียของเซลล์พืชและสัตว์ และเฉพาะเมื่อมีออกซิเจนเท่านั้น นี่คือชุดของการเปลี่ยนแปลงทางเคมีก่อนการก่อตัวของผลิตภัณฑ์ขั้นสุดท้าย - คาร์บอนไดออกไซด์ ในขั้นตอนต่างๆ ของกระบวนการนี้ ผลิตภัณฑ์ขั้นกลางของการเกิดออกซิเดชันของสารตั้งต้นจะเกิดขึ้นพร้อมกับการกำจัดอะตอมของไฮโดรเจน ในกรณีนี้พลังงานจะถูกปล่อยออกมาซึ่ง "อนุรักษ์" ในพันธะเคมีของ ATP และโมเลกุลของน้ำจะเกิดขึ้น เห็นได้ชัดว่าจำเป็นต้องจับอะตอมไฮโดรเจนที่แยกออกจากกันซึ่งจำเป็นต้องใช้ออกซิเจนอย่างแม่นยำ การเปลี่ยนแปลงทางเคมีชุดนี้ค่อนข้างซับซ้อนและเกิดขึ้นจากการมีส่วนร่วมของเยื่อหุ้มภายในของไมโตคอนเดรีย เอนไซม์ และโปรตีนพาหะ
การหายใจระดับเซลล์มีประสิทธิภาพมาก มีการสังเคราะห์โมเลกุล ATP 30 โมเลกุล อีกสองโมเลกุลจะเกิดขึ้นระหว่างไกลโคไลซิส และโมเลกุลเอทีพีหกโมเลกุลเกิดขึ้นอันเป็นผลมาจากการเปลี่ยนแปลงของผลิตภัณฑ์ไกลโคไลซิสบนเยื่อหุ้มไมโตคอนเดรีย โดยรวมแล้วอันเป็นผลมาจากการเกิดออกซิเดชันของโมเลกุลกลูโคสหนึ่งโมเลกุลทำให้เกิดโมเลกุล ATP 38 โมเลกุล:
ค 6 ชม. 12 โอ 6 + 6H 2 0 → 6CO 2 + 6H 2 O + 38ATP
ขั้นตอนสุดท้ายของการเกิดออกซิเดชันไม่เพียงแต่น้ำตาลเท่านั้น แต่ยังรวมถึงโปรตีนและไขมันในไมโตคอนเดรียด้วย สารเหล่านี้ถูกใช้โดยเซลล์ โดยส่วนใหญ่เมื่อปริมาณคาร์โบไฮเดรตหมดลง ขั้นแรกให้บริโภคไขมันซึ่งออกซิเดชั่นจะปล่อยพลังงานออกมามากกว่าคาร์โบไฮเดรตและโปรตีนในปริมาณที่เท่ากัน ดังนั้นไขมันในสัตว์จึงถือเป็น "การสำรองทางยุทธศาสตร์" หลักของแหล่งพลังงาน ในพืช แป้งมีบทบาทในการสำรองพลังงาน เมื่อเก็บไว้ จะใช้พื้นที่มากกว่าปริมาณไขมันเทียบเท่าพลังงานอย่างมาก นี่ไม่ใช่อุปสรรคสำหรับพืชเนื่องจากพวกมันไม่สามารถเคลื่อนที่ได้และไม่ได้ขนเสบียงติดตัวเหมือนกับสัตว์ คุณสามารถดึงพลังงานจากคาร์โบไฮเดรตได้เร็วกว่าจากไขมันมาก โปรตีนทำหน้าที่สำคัญมากมายในร่างกาย ดังนั้นจึงเกี่ยวข้องกับการเผาผลาญพลังงานเฉพาะเมื่อทรัพยากรของน้ำตาลและไขมันหมดลง เช่น ในระหว่างการอดอาหารเป็นเวลานาน
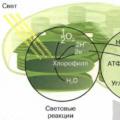
การสังเคราะห์แสง. การสังเคราะห์ด้วยแสงเป็นกระบวนการที่พลังงานของรังสีดวงอาทิตย์ถูกแปลงเป็นพลังงานของพันธะเคมีของสารประกอบอินทรีย์ ในเซลล์พืช กระบวนการที่เกี่ยวข้องกับการสังเคราะห์ด้วยแสงเกิดขึ้นในคลอโรพลาสต์ ภายในออร์แกเนลล์นี้มีระบบเมมเบรนซึ่งมีเม็ดสีฝังอยู่เพื่อจับพลังงานรังสีของดวงอาทิตย์ เม็ดสีหลักของการสังเคราะห์ด้วยแสงคือคลอโรฟิลล์ ซึ่งดูดซับสีน้ำเงินและสีม่วงเป็นส่วนใหญ่ รวมถึงรังสีสีแดงของสเปกตรัม แสงสีเขียวจะสะท้อนออกมา ดังนั้นคลอโรฟิลล์เองและส่วนต่างๆ ของพืชที่ประกอบด้วยคลอโรฟิลล์จึงปรากฏเป็นสีเขียว
การสังเคราะห์ด้วยแสงมี 2 ระยะ คือ แสงสว่างและ มืด(รูปที่ 53) การดักจับและการแปลงพลังงานรังสีที่เกิดขึ้นจริงเกิดขึ้นในช่วงระยะแสง เมื่อดูดซับควอนตัมแสง คลอโรฟิลล์จะเข้าสู่สภาวะตื่นเต้นและกลายเป็นผู้บริจาคอิเล็กตรอน อิเล็กตรอนของมันถูกถ่ายโอนจากโปรตีนคอมเพล็กซ์หนึ่งไปยังอีกโปรตีนหนึ่งตามห่วงโซ่การขนส่งอิเล็กตรอน โปรตีนของสายโซ่นี้ เช่นเดียวกับเม็ดสี จะเข้มข้นอยู่ที่เยื่อหุ้มชั้นในของคลอโรพลาสต์ เมื่ออิเล็กตรอนเคลื่อนที่ไปตามสายโซ่พาหะ จะสูญเสียพลังงานซึ่งใช้สำหรับการสังเคราะห์ ATP อิเล็กตรอนบางตัวที่ถูกกระตุ้นด้วยแสงถูกใช้เพื่อลด NDP (nicotinamide adenine dinucleotiphosphate) หรือ NADPH

ข้าว. 53. ผลปฏิกิริยาของระยะแสงและความมืดของการสังเคราะห์ด้วยแสง
ภายใต้อิทธิพลของแสงแดด โมเลกุลของน้ำก็ถูกสลายเป็นคลอโรพลาสต์เช่นกัน - โฟโตไลซิส; ในกรณีนี้อิเล็กตรอนจะปรากฏขึ้นเพื่อชดเชยการสูญเสียคลอโรฟิลล์ สิ่งนี้จะผลิตออกซิเจนเป็นผลพลอยได้:
ดังนั้นความหมายเชิงหน้าที่ของเฟสแสงคือการสังเคราะห์ ATP และ NADPH โดยการแปลงพลังงานแสงเป็นพลังงานเคมี
แสงไม่จำเป็นสำหรับการสังเคราะห์ด้วยแสงในช่วงมืด สาระสำคัญของกระบวนการที่เกิดขึ้นที่นี่คือโมเลกุล ATP และ NADPH ที่ผลิตในช่วงแสงจะถูกใช้ในปฏิกิริยาเคมีชุดหนึ่งที่ "แก้ไข" CO2 ในรูปของคาร์โบไฮเดรต ปฏิกิริยาเฟสมืดทั้งหมดเกิดขึ้นภายในคลอโรพลาสต์ และคาร์บอนไดออกไซด์ ADP และ NADP ที่ปล่อยออกมาระหว่าง "การตรึง" จะถูกนำมาใช้อีกครั้งในปฏิกิริยาเฟสแสงสำหรับการสังเคราะห์ ATP และ NADPH
สมการโดยรวมสำหรับการสังเคราะห์ด้วยแสงมีดังนี้:
ความสัมพันธ์และความสามัคคีของกระบวนการแลกเปลี่ยนพลาสติกและพลังงาน. กระบวนการสังเคราะห์ ATP เกิดขึ้นในไซโตพลาสซึม (ไกลโคไลซิส) ในไมโตคอนเดรีย (การหายใจของเซลล์) และในคลอโรพลาสต์ (การสังเคราะห์ด้วยแสง) ปฏิกิริยาทั้งหมดที่เกิดขึ้นระหว่างกระบวนการเหล่านี้เป็นปฏิกิริยาการแลกเปลี่ยนพลังงาน พลังงานที่เก็บไว้ในรูปของ ATP จะถูกใช้ในปฏิกิริยาการแลกเปลี่ยนพลาสติกเพื่อผลิตโปรตีน ไขมัน คาร์โบไฮเดรต และกรดนิวคลีอิกที่จำเป็นสำหรับชีวิตของเซลล์ โปรดทราบว่าระยะมืดของการสังเคราะห์ด้วยแสงนั้นเป็นลูกโซ่ของปฏิกิริยา การแลกเปลี่ยนพลาสติก และระยะแสงคือการแลกเปลี่ยนพลังงาน
ความสัมพันธ์และความสามัคคีของกระบวนการพลังงานและการแลกเปลี่ยนพลาสติกแสดงให้เห็นได้ดีจากสมการต่อไปนี้:

เมื่ออ่านสมการนี้จากซ้ายไปขวา เราจะได้กระบวนการออกซิเดชันของกลูโคสไปเป็นคาร์บอนไดออกไซด์และน้ำระหว่างไกลโคไลซิสและการหายใจของเซลล์ ซึ่งเกี่ยวข้องกับการสังเคราะห์ ATP (การเผาผลาญพลังงาน) หากคุณอ่านจากขวาไปซ้าย คุณจะได้คำอธิบายเกี่ยวกับปฏิกิริยาในช่วงมืดของการสังเคราะห์ด้วยแสง เมื่อกลูโคสถูกสังเคราะห์จากน้ำและคาร์บอนไดออกไซด์โดยมีส่วนร่วมของ ATP (การแลกเปลี่ยนพลาสติก)
การสังเคราะห์ทางเคมี. นอกจากโฟโตออโตโทรฟแล้ว แบคทีเรียบางชนิด (แบคทีเรียไฮโดรเจน แบคทีเรียไนตริไฟนิ่ง แบคทีเรียซัลเฟอร์ ฯลฯ) ยังสามารถสังเคราะห์สารอินทรีย์จากสารอนินทรีย์ได้อีกด้วย พวกมันทำการสังเคราะห์นี้เนื่องจากพลังงานที่ปล่อยออกมาระหว่างการออกซิเดชั่นของสารอนินทรีย์ พวกมันถูกเรียกว่าคีโมออโตโทรฟ แบคทีเรียสังเคราะห์ทางเคมีเหล่านี้มีบทบาทสำคัญในชีวมณฑล ตัวอย่างเช่น แบคทีเรียไนตริไฟริ่งจะเปลี่ยนเกลือแอมโมเนียมที่ไม่สามารถดูดซึมโดยพืชไปเป็นเกลือของกรดไนตริกซึ่งพวกมันดูดซึมได้ดี
เมแทบอลิซึมของเซลล์ประกอบด้วยปฏิกิริยาของพลังงานและเมแทบอลิซึมของพลาสติก ในระหว่างการเผาผลาญพลังงาน สารประกอบอินทรีย์ที่มีพันธะเคมีพลังงานสูง - ATP - จะเกิดขึ้น พลังงานที่จำเป็นสำหรับสิ่งนี้มาจากการออกซิเดชันของสารประกอบอินทรีย์ในระหว่างปฏิกิริยาไม่ใช้ออกซิเจน (ไกลโคไลซิส การหมัก) และแอโรบิก (การหายใจของเซลล์) จากแสงแดดพลังงานที่ถูกดูดซับในช่วงแสง (การสังเคราะห์ด้วยแสง) จากปฏิกิริยาออกซิเดชันของสารประกอบอนินทรีย์ (การสังเคราะห์ทางเคมี) พลังงาน ATP ถูกใช้ไปกับการสังเคราะห์สารประกอบอินทรีย์ที่จำเป็นสำหรับเซลล์ในระหว่างปฏิกิริยาการแลกเปลี่ยนพลาสติก ซึ่งรวมถึงปฏิกิริยาในช่วงมืดของการสังเคราะห์ด้วยแสง
- อะไรคือความแตกต่างระหว่างการเผาผลาญพลาสติกและพลังงาน?
- พลังงานของแสงอาทิตย์ถูกแปลงเป็นระยะแสงของการสังเคราะห์ด้วยแสงอย่างไร? กระบวนการใดเกิดขึ้นในช่วงมืดของการสังเคราะห์ด้วยแสง?
- เหตุใดการสังเคราะห์ด้วยแสงจึงเรียกว่ากระบวนการสะท้อนปฏิสัมพันธ์ระหว่างดาวเคราะห์และจักรวาล