Embrionalni razvoj je složen i dugotrajan morfogenetski proces tijekom kojeg se iz očevih i majčinih zametnih stanica formira novi višestanični organizam, sposoban za samostalan život u uvjetima okoliša. U osnovi je spolnog razmnožavanja i osigurava prijenos nasljednih karakteristika s roditelja na potomstvo.
Oplodnja se sastoji od spajanja spermija s jajnom stanicom. Najvažnije faze procesa oplodnje su:
1) prodiranje SP u jaje;
2) aktivacija raznih sintetskih procesa u jajetu;
3) fuzija jezgre jajeta i SP s obnavljanjem diploidnog skupa kromosoma.
Da bi došlo do oplodnje, ženske i muške spolne stanice moraju se spojiti. To se postiže oplodnjom.
Prodor SP u jajnu stanicu pospješuju enzim hijaluronidijaza i druge biološki aktivne tvari (spermolizin) koje povećavaju propusnost glavne međustanične tvari.Enzime luči akrosom tijekom procesa akrosomska reakcija. Njegova bit je sljedeća: u trenutku kontakta s jajnom stanicom na vrhu glave spermija, plazma membrana i membrana uz nju se otapaju, a susjedni dio jajne membrane se otapa.Akrosomska membrana strši prema van i oblikuje izraslina u obliku šuplje cijevi. U području takvog kontakta nastaje izbočina ili oplodni tuberkul. Nakon čega se spajaju plazma membrane obiju spolnih stanica i počinje sjedinjavanje njihovog sadržaja. Od tog trenutka zajednički pothvat a I predstavlja jednu stanicu zigote.
Aktivacija I ili kortikalna reakcija, koji se razvija kao rezultat kontakta sa SP, ima morfološke i biokemijske manifestacije. Aktivacija se očituje promjenama u površinskom kortikalnom sloju ooplazme i tvorbi oplodne opne. Oplodna membrana štiti jajnu stanicu od prodiranja oplođenih spermija.
zigota- diploidna (sadrži kompletan dvostruki skup kromosoma) stanica nastala kao rezultat oplodnje (fuzije jajne stanice i spermija). Zigota je totipotentan(odnosno, sposobna stvoriti bilo koju drugu) stanicu.
U čovjeka se prva mitotička dioba zigote događa otprilike 30 sati nakon oplodnje, što je posljedica složenih procesa pripreme za prvu diobu drobljenje.
Razdvajanje - ovo je niz uzastopnih mitotskih dioba zigote koje završavaju stvaranjem višestaničnog embrija - blastule. Prva cijepna dioba počinje nakon sjedinjenja nasljednog materijala pronukleusa i stvaranja zajedničke metafazne ploče. Stanice koje nastaju tijekom cijepanja nazivaju se blastomeri(od grčkog eksplodirati- klica, klica). Značajka mitotičkih dioba je da sa svakom diobom stanice postaju sve manje i manje sve dok ne dostignu omjer volumena jezgre i citoplazme koji je uobičajen za somatske stanice.Prvo, blastomeri se nalaze jedan uz drugoga, tvoreći klaster stanice tzv Morula . Tada se između stanica formira šupljina - blastocel , ispunjen tekućinom. Stanice su potisnute na periferiju, tvoreći stijenku blastule - blastoderm. Ukupna veličina embrija na kraju cijepanja u fazi blastule ne prelazi veličinu zigote.
Progeneza – gametogeneza (spermato- i oogeneza) i oplodnja Spermatogeneza se odvija u zavijenim tubulima testisa i dijeli se na četiri razdoblja: 1) reproduktivno razdoblje – I; 2) razdoblje rasta – II; 3) rok dozrijevanja – III; 4) razdoblje formiranja – IV. Oogeneza se odvija u jajnicima i dijeli se na tri razdoblja: 1) reproduktivno razdoblje (u embriogenezi i tijekom prve godine nakon embrionalni razvoj); 2) razdoblje rasta (mali i veliki); 3) razdoblje sazrijevanja Jaje se sastoji od jezgre s haploidnim skupom kromosoma i izraženom citoplazmom, koja sadrži sve organele, s izuzetkom citocentra.
Razdvajanje. Karakteristike drobljenja. Glavne vrste jaja prema položaju žumanjka. Povezanost građe jajeta s vrstom cijepanja. Blastomeri i embrionalne stanice. Građa i vrste blastula.
Razdvajanje - Ovaj se proces temelji na mitotičkoj diobi stanica. Međutim, stanice kćeri nastale kao rezultat diobe ne raspršuju se, već ostaju bliske jedna uz drugu. Tijekom procesa cijepanja, stanice kćeri postupno postaju manje. Svaku životinju karakterizira određena vrsta drobljenja, određena količinom i prirodom raspodjele žumanjka u jajetu. Žumanjak sprječava drobljenje, stoga se dio zigote preopterećen fragmentima žumanjka sporije ili se uopće ne fragmentira.
U izolecitalnom, žumanjkom siromašno oplođeno kopljasto jaje, prva cijepna brazda u obliku proreza počinje na animalnom polu i postupno se širi u uzdužnom meridionalnom smjeru do vegetativnog, dijeleći jaje na 2 stanice - 2 blastomera. Druga brazda ide okomito na prvu - nastaju 4 blastomera. Kao rezultat niza uzastopnih fragmentacija, formiraju se skupine stanica blisko jedna uz drugu. Kod nekih životinja takav embrij podsjeća na dud ili malinu. Dobio je ime morule(latinski morum - dud) - višestanična lopta bez šupljine iznutra.
U telolecitalna jaja , preopterećeno žumanjkom – drobljenje može biti potpuno jednolično ili neravnomjerno i nepotpuno. Zbog obilja inertnog žumanjka, blastomeri vegetativnog pola uvijek zaostaju u brzini fragmentacije za blastomerima animalnog pola. Za jaja vodozemaca karakteristično je potpuno, ali neravnomjerno drobljenje. Kod riba, ptica i nekih drugih životinja zgnječen je samo dio jajeta koji se nalazi na animalnom polu; dolazi do nepotpune diskoidne fragmentacije. Tijekom procesa cijepanja povećava se broj blastomera, ali blastomeri ne rastu do veličine izvorne stanice, već svakim cijepanjem postaju sve manji. To se objašnjava činjenicom da mitotski ciklusi zigote cijepanja nemaju tipičnu interfazu; presintetsko razdoblje (G1) je odsutno, a sintetsko razdoblje (S) počinje u telofazi prethodne mitoze.
Gnječenje jaja završava formiranjem blastule.
U polilecitalnim jajima ribe koštunjače, gmazovi, ptice, kao i monotreme sisavci drobljenje djelomično, ili merob-lastic, oni. pokriva samo citoplazmu bez žumanjka. Nalazi se u obliku tankog diska na animalnom polu, stoga ovaj tip drobljenje se zove diskoidan . Pri karakterizaciji vrste fragmentacije također se uzimaju u obzir relativni položaj i brzina diobe blastomera. Ako su blastomeri poredani u redove jedan iznad drugog duž radijusa, drobljenje se zove radijalno.
Fragmentacija može biti: deterministička i regulatorna; potpuni (holoblastični) ili nepotpuni (meroblastični); jednoliki (blastomeri su više-manje iste veličine) i nejednaki (blastomeri nisu iste veličine, razlikuju se dvije ili tri skupine veličina, obično se nazivaju makro- i mikromeri)
Vrste jaja:
Količina žumanjka - Oligolecetal (lancelet) Mezolecetal (vodozemci) Polilecetal (ribe, ptice)
Mjesto- Izolecetalni(smješten difuzno, ravnomjerno). Sadrže malo žumanjka, ravnomjerno raspoređenog po stanici. Karakteristično za bodljikaše, niže hordate i sisavce. Kod sisavaca to su alecijalna jaja (praktički nema žumanjka)
Telolecetal(s umjerenim količinama žumanjka na donjem vegetativnom polu)
Jako telolecitalan (s velikom količinom žumanjka, zauzima cijelo jaje osim gornjeg pola. Ima dosta žumanjka, koncentriran na vegetativnom polu. Postoje 2 skupine: umjereno telolecitan (mekušci, vodozemci) i oštro lecitalan (gmazovi). i ptice).Citoplazma i jezgra su koncentrirani na anomalnom polu.
Centrolecetalni(ima malo žumanjka, ali čvrsto u sredini). U sredini je malo žumanjka. Karakteristično za člankonošce
Blastomeri- stanice nastale kao rezultat diobe jaja u višestaničnih životinja. Karakteristična značajka B. je odsutnost rasta u razdoblju između podjela, zbog čega se sa sljedećom podjelom volumen svakog B. prepolovljuje. S holoblastikom Kada se zgnječe u telolecitalnim jajima, B. se razlikuju po veličini: veliki B. su makromeri, srednji su mezomeri, mali su mikromeri. Tijekom sinkronih dioba, fragmentacija bakterija je u pravilu jednoličnog oblika, a struktura njihove citoplazme je vrlo jednostavna. Tada se površinske bakterije spljošte, a jaje prelazi u završnu, fazu drobljenja - blastulacija.
Građa blastule. Ako se formira čvrsta lopta bez unutarnje šupljine, tada se takav embrij naziva Morula. Nastanak blastule ili morule ovisi o svojstvima citoplazme. Blastula nastaje kada je citoplazma dovoljno viskozna, a morula kada je viskoznost slaba. S dovoljnom viskoznošću citoplazme, blastomeri zadržavaju zaobljeni oblik i blago su spljošteni samo na mjestima kontakta. Kao rezultat toga, između njih se pojavljuje jaz, koji se, kako se fragmentira, povećava, puni tekućinom i pretvara u blastocoel. Kada je viskoznost citoplazme slaba, blastomeri nisu zaobljeni i nalaze se blizu jedan uz drugi, nema razmaka i ne stvara se šupljina. Blastule se razlikuju po strukturi i ovise o vrsti cijepanja.
Začetak novog organizma daje oplođeno jajašce (osim slučajeva partenogeneze i vegetativnog razmnožavanja). Oplodnja je proces međusobnog spajanja dviju spolnih stanica (gameta) pri čemu se obavljaju dvije različite funkcije: spolna (spajanje gena dvaju roditelja) i reproduktivna (nastanak novog organizma). Prva od ovih funkcija uključuje prijenos gena s roditelja na potomstvo, druga je inicijacija u citoplazmi jajašca onih reakcija i pokreta koji omogućuju nastavak razvoja. Kao rezultat oplodnje, dvostruki (2n) set kromosoma se obnavlja u jajetu. Centrosom koji unese spermij nakon duplikacije formira fisijsko vreteno, a zigota ulazi u 1. fazu embriogeneze – fazu cijepanja. Kao rezultat mitoze, iz zigote nastaju 2 stanice kćeri - blastomeri.
Prezigotsko razdoblje
Prezigotsko razdoblje razvoja povezano je sa stvaranjem gameta (gametogeneza). Stvaranje jajnih stanica počinje kod žena i prije njihova rođenja, a završava se za svako dano jaje tek nakon njegove oplodnje. Ženski fetus do rođenja u jajnicima sadrži oko dva milijuna oocita prvog reda (to su još uvijek diploidne stanice), a samo 350 - 450 njih će doći u stadij oocita drugog reda (haploidne stanice) , pretvarajući se u jaja (jedno po jedno tijekom jednog menstrualnog ciklusa). Za razliku od žena, spolne stanice u testisima (testisi) kod muškaraca počinju se formirati tek s početkom puberteta. Trajanje stvaranja spermija je otprilike 70 dana; po gramu težine testisa, broj spermija je oko 100 milijuna dnevno.
Gnojidba
Gnojidba - spajanje muške spolne stanice (spermija) sa ženskom (jaje, jajašce), što dovodi do stvaranja zigote – novog jednostaničnog organizma. Biološki smisao oplodnje je sjedinjavanje nuklearnog materijala muških i ženskih spolnih stanica, što dovodi do sjedinjavanja očevih i majčinih gena, obnavljanja diploidnog sklopa kromosoma, kao i aktivacije jajne stanice, odnosno stimulacije njegov embrionalni razvoj. Spajanje jajne stanice sa spermijem obično se događa u proširenom dijelu jajovoda u obliku lijevka tijekom prvih 12 sati nakon ovulacije.
Sjemena tekućina, koja tijekom spolnog odnosa ulazi u vaginu žene, obično sadrži od 60 do 150 milijuna spermija, koji zahvaljujući pokretima brzinom od 2-3 mm u minuti, stalnim valovitim kontrakcijama maternice i jajovoda te alkalnoj sredini , već nakon 1-2 minute nakon spolnog odnosa dospijevaju u maternicu, a nakon 2-3 sata - krajnje dijelove jajovoda, gdje se obično događa spajanje s jajetom. Postoje monospermični (jedan spermij prodire u jaje) i polispermij (dva ili više spermija prodiru u jaje, ali se samo jedna jezgra spermija spaja s jezgrom jajeta). Očuvanje aktivnosti spermija tijekom prolaska kroz genitalni trakt žene olakšava blago alkalno okruženje cervikalnog kanala maternice, ispunjenog čepom sluzi. Tijekom orgazma tijekom spolnog odnosa sluzni čep iz cervikalnog kanala se djelomično istisne, a potom ponovno uvuče u njega i na taj način omogućuje brži ulazak sperme iz rodnice (gdje inače zdrava žena blago kisela sredina) u povoljniju okolinu vrata maternice i šupljine maternice. Prolaz sperme kroz sluzni čep cervikalnog kanala također je olakšan naglo povećanom propusnošću sluzi u danima ovulacije. U preostalim danima menstrualnog ciklusa sluzni čep ima znatno manju propusnost za spermu.
Mnogi spermiji koji se nalaze u ženinom genitalnom traktu mogu zadržati sposobnost oplodnje 48-72 sata (ponekad i do 4-5 dana). Ovulirano jaje ostaje održivo otprilike 24 sata. Uzimajući to u obzir, najpovoljnije vrijeme za oplodnju smatra se razdoblje rupture zrelog folikula nakon čega slijedi rođenje jajašca, kao i 2-3 dana nakon ovulacije. Žene koje koriste fiziološku metodu kontracepcije trebaju imati na umu da vrijeme ovulacije može varirati, a održivost jajne stanice i spermija može biti znatno dulja. Ubrzo nakon oplodnje, zigota se počinje fragmentirati i formirati embrij.
zigotaZigota (grč. zigota u paru) je diploidna (koja sadrži kompletan dvostruki skup kromosoma) stanica nastala kao rezultat oplodnje (spajanja jajne stanice i spermija). Zigota je totipotentna (to jest, sposobna rađati bilo koju drugu) stanica. Pojam je uveo njemački botaničar E. Strassburger.
U čovjeka se prva mitotička dioba zigote događa otprilike 30 sati nakon oplodnje, što je posljedica složenih procesa pripreme za prvi čin cijepanja. Stanice nastale kao rezultat fragmentacije zigote nazivaju se blastomere. Prve diobe zigote nazivaju se "fragmentacije" jer je stanica fragmentirana: stanice kćeri postaju manje nakon svake diobe, a između dioba nema stadija rasta stanice.
Razvoj zigote Zigota se ili počinje razvijati odmah nakon oplodnje, ili je prekrivena gustom ljuskom i neko vrijeme se pretvara u sporu koja miruje (često se naziva zigospora) - karakteristična za mnoge gljive i alge.
RazdvajanjeRazdoblje embrionalnog razvoja višestanične životinje počinje fragmentacijom zigote i završava rođenjem nove jedinke. Proces cijepanja sastoji se od niza uzastopnih mitotskih dioba zigote. Dvije stanice nastale kao rezultat nove diobe zigote i svih sljedećih generacija stanica u ovoj fazi nazivaju se blastomere. Tijekom fragmentacije jedna dioba slijedi drugu, a nastali blastomeri ne rastu, zbog čega je svaka nova generacija blastomera predstavljena manjim stanicama. Ova značajka diobe stanica tijekom razvoja oplođenog jajašca odredila je pojavu figurativnog izraza - fragmentacije zigote.
U različiti tipoviživotinjska se jaja razlikuju po količini i naravi raspodjele rezervnih hranjivih tvari (žumanjka) u citoplazmi. To uvelike određuje prirodu naknadne fragmentacije zigote. S malom količinom i ravnomjernom raspodjelom žumanjka u citoplazmi, cijela masa zigote se dijeli uz stvaranje identičnih blastomera - potpuna ravnomjerna fragmentacija (na primjer, kod sisavaca). Kada se žumanjak nakuplja pretežno na jednom od polova zigote, dolazi do neravnomjerne fragmentacije - nastaju blastomeri koji se razlikuju po veličini: veći makromeri i mikromeri (na primjer, u vodozemaca). Ako je jaje jako bogato žumanjkom, tada se dio bez žumanjka zdrobi. Dakle, kod gmazova i ptica samo je diskasti dio zigote na jednom od polova, gdje se nalazi jezgra, podložan fragmentaciji - nepotpunoj, diskoidnoj fragmentaciji. Konačno, kod insekata samo površinski sloj citoplazme zigote sudjeluje u procesu drobljenja – nepotpuno, površinsko drobljenje.
Kao rezultat fragmentacije (kada broj dijeljenih blastomera dosegne značajan broj), nastaje blastula. U tipičnom slučaju (na primjer, u lanceletu), blastula je šuplja lopta, čiju stijenku čini jedan sloj stanica (blastoderm). Šupljina blastule - blastocoel, inače nazvana primarna tjelesna šupljina, ispunjena je tekućinom. Kod vodozemaca blastula ima vrlo malu šupljinu, a kod nekih životinja (na primjer, člankonožaca) blastocel može biti potpuno odsutan.
GastrulacijaU sljedećoj fazi embrionalnog razdoblja dolazi do procesa formiranja gastrule - gastrulacije. Kod mnogih životinja stvaranje gastrule događa se invaginacijom, tj. izbočenje blastoderma na jednom od polova blastule (s intenzivnom proliferacijom stanica u ovoj zoni). Kao rezultat toga, formira se dvoslojni embrij u obliku šalice. Vanjski sloj stanica je ektoderm, a unutarnji sloj je endoderm. Unutarnja šupljina koja nastaje izbočenjem stijenke blastule, primarno crijevo, komunicira s vanjskom okolinom kroz otvor - primarna usta (blastopore). Postoje i druge vrste gastrulacije. Na primjer, kod nekih koelenterata endoderm gastrule nastaje imigracijom, tj. "izbacivanje" nekih stanica blastoderme u šupljinu blastule i njihovo naknadno razmnožavanje. Primarna usta nastaju rupturom stijenke gastrule. Uz neravnomjernu fragmentaciju (kod nekih crva i mekušaca), gastrula nastaje kao rezultat rasta makromera s mikromerima i stvaranja endoderma na račun prvog. Često se kombiniraju različite metode gastrulacije.
U svih životinja (osim spužvi i koelenterata - dvoslojnih životinja) stadij gastrulacije završava stvaranjem drugog sloja stanica - mezoderma. Ovaj "stanični sloj nastaje između ento- i ektoderma. Postoje dva poznata načina polaganja mezoderma. Kod anelida, na primjer, u području gastrule blastopore, odvajaju se dvije velike stanice (teloblasti). Prilikom razmnožavanja, iz njih nastaju dvije mezodermalne pruge, od kojih (dijelom zbog divergencije stanica, dijelom kao rezultat razaranja dijela stanica unutar mezodermalnih pruga) nastaju celomske vrećice - teloblastična metoda polaganja mezoderma. enterokoelnoj metodi (bodljikaši, lancetaši, kralješnjaci), kao posljedica izbočenja stijenke primarnog crijeva nastaju bočni džepovi koji se potom odvajaju i postaju celomske vrećice.U oba slučaja polaganja mezoderma, celomske vrećice rastu i ispunjavaju primarna tjelesna šupljina.Mezodermalni sloj stanica koji oblaže tjelesnu šupljinu iznutra tvori peritonealni epitel.Šupljina, koja zamjenjuje primarnu, naziva se sekundarna tjelesna šupljina ili coelom.U slučaju teloblastične metode polaganja mezoderma blastopora se pretvara u usni otvor odrasle životinje. Takvi se organizmi nazivaju protostomi. Kod deuterostomnih životinja (s enterokoelnom metodom polaganja mezoderma) blastopora prerasta ili se pretvara u anus, a usta odrasle jedinke pojavljuju se drugi put, izbočenjem ektoderma.
Formiranje triju klica (ekto-, ento- i mezoderma) završava stadij gastrulacije, a od tog trenutka počinju procesi histo- i organogeneze. Kao rezultat diferencijacije stanica triju klica nastaju različita tkiva i organi organizma u razvoju. Još krajem prošlog stoljeća (uglavnom zahvaljujući istraživanjima I. I. Mečnikova i A. O. Kovalevskog) utvrđeno je da kod različitih životinjskih vrsta isti klicini listići daju iste organe i tkiva. Iz ektoderma nastaju epidermis sa svim njegovim izvedenim strukturama i živčani sustav. Endoderm tvori probavni trakt i pripadajuće organe (jetra, gušterača, pluća itd.). Mezoderm tvori kostur, krvožilni sustav, ekskretorni aparat i spolne žlijezde. Iako se danas klice ne smatraju strogo specijaliziranim, njihova je homologija kod velike većine životinjskih vrsta očita, što ukazuje na jedinstvo podrijetla životinjskog carstva.
Tijekom embrionalnog razdoblja povećava se stopa rasta i diferencijacije organizama u razvoju. Ako tijekom procesa fragmentacije ne dođe do rasta i blastula (u svojoj masi) može biti znatno inferiorna u odnosu na zigotu, tada se, počevši od procesa gastrulacije, masa embrija brzo povećava (zbog intenzivne reprodukcije stanica). Procesi stanične diferencijacije započinju u najranijoj fazi embriogeneze – fragmentacija i u podlozi su primarne diferencijacije tkiva – nastanak triju klica (embrionalna tkiva). Daljnji razvoj embrija prati sve intenzivniji proces diferencijacije tkiva i organa. Kao rezultat embrionalnog razdoblja razvoja, formira se organizam koji je sposoban samostalno (manje ili više) postojati u vanjskom okruženju. Rođenje nove jedinke događa se ili kao rezultat izlijeganja iz jajeta (kod jajorodnih životinja) ili napuštanja majčinog tijela (kod viviparnih životinja).
Histo- i organogenezaHisto- i organogeneza embrija odvija se kao rezultat reprodukcije, migracije, diferencijacije stanica i njihovih sastavnih dijelova, uspostavljanja međustaničnih kontakata i smrti nekih stanica. Presomitno razdoblje nastavlja se od 317. do 20. dana; somitno razdoblje razvoja počinje 20. dana. Dvadesetog dana embriogeneze, stvaranjem nabora trupa (cefalokaudalnih i bočnih), dolazi do odvajanja samog embrija od ekstraembrionalnih organa, kao i promjene njegovog ravnog oblika u cilindrični. U isto vrijeme, dorzalni dijelovi mezoderma embrija podijeljeni su u zasebne segmente koji se nalaze s obje strane notohorde - somite. 21. dana embrij ima 2-3 para somita. Somiti se počinju formirati od trećeg para, a prvi i drugi par pojavljuju se nešto kasnije. Broj somita postupno raste: 23. dana razvoja ima 10 pari somita, 25. - 14 parova, 27. - 25 parova, na kraju petog tjedna broj somita u embriju doseže 43. -44 para. Na temelju brojanja somita moguće je približno odrediti vrijeme razvoja (somitičku dob) embrija.
Iz vanjskog dijela svakog somita nastaje dermatom, iz unutarnjeg dijela sklerotom, a iz srednjeg dijela miotom. Dermatom postaje izvorište dermisa kože, sklerotoma - hrskavice i koštanog tkiva, miotoma - skeletnih mišića dorzalnog dijela embrija. Ventralni dijelovi mezoderma - splanhnotom - nisu segmentirani, već su podijeljeni na visceralni i parijetalni sloj, iz kojih se razvijaju serozne membrane unutarnjih organa, mišića srca i kore nadbubrežne žlijezde. Iz mezenhima splanhnotoma nastaju krvne žile, krvne stanice, vezivno i glatko mišićno tkivo embrija. Područje mezoderma koje povezuje somite sa splanhnotomom podijeljeno je na segmentne noge - nefrogonotome, koje služe kao izvor razvoja bubrega i spolnih žlijezda, kao i paramezonefricnih kanala. Potonji tvori epitel maternice i jajovoda.
Tijekom procesa diferencijacije embrionalnog ektoderma nastaju neuralna cijev, neuralni grebeni, plakode, kožni ektoderm i prehordalna ploča. Proces formiranja neuralne cijevi naziva se neurulacija. Sastoji se od stvaranja udubljenja poput proreza na površini ektoderma; zadebljani rubovi ovog udubljenja (živčani nabori) srastaju i tvore neuralnu cijev. Iz kranijalnog dijela neuralne cijevi nastaju moždani mjehurići - rudiment mozga. S obje strane neuralne cijevi (između potonje i kožnog ektoderma) odvajaju se skupine stanica od kojih nastaju neuralni grebeni. Stanice neuralnog grebena sposobne su za migraciju. Stanice koje migriraju prema dermatomu daju pigmentne stanice – melanocite; Stanice neuralnog grebena koje migriraju prema trbušnoj šupljini stvaraju simpatičke i parasimpatičke ganglije, srž nadbubrežne žlijezde. Od stanica neuralnog grebena koje ne migriraju nastaju ganglijske ploče iz kojih se razvijaju spinalni i periferni autonomni živčani gangliji. Plakode tvore ganglije glave i živčane stanice organa sluha i ravnoteže.
Razdvajanje(segmentacija) u pojedinih predstavnika razreda kralježnjaka ima općenito isti tijek; međutim, kao što je već spomenuto, na njega su utjecali čimbenici koji su tijekom filogeneze utjecali na razvoj u obliku posljedica utjecaja unutarnje i vanjske sredine u kojoj su organizmi živjeli tijekom svog predačkog razvoja (cenogenetski čimbenici).
Prilikom promatranja promjene, koji se javljaju u jajima prema filogenetskom razvoju jaja pojedinih predstavnika razreda kralješnjaka, može se primijetiti da se jajne stanice međusobno značajno razlikuju po sadržaju hranjive i gradivne tvari - žumanjka. Jajne stanice kopljača (Amfioxus), organizma koji se filogenetski smatra najniže organiziranim bićem, ali koji već ima jaku dorzalnu regiju, svrstavaju se u oligolecitale.
Međutim, u skladu s filogenetskim razvojem, količina žumanjka u jajima kralježnjaka, koji su filogenetski najorganiziraniji organizmi, sve više raste, a maksimalnu količinu dostiže u jajima ptica, koja su relativno velika i polilecitalna. Pod utjecajem cenogenetskih čimbenika (čimbenika koji djeluju iz vanjske okoline i uzrokovanih promjenama u načinu života, a samim time i razvoja), količina žumanjka u procesu filogenetskog razvoja prema čovjeku sve se više smanjuje, zbog čega jajašca čovjeka i viši sisavci ponovno postaju (sekundarno) oligolecitalni .
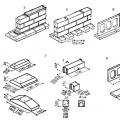
Prisutnost varijabilnog iznosa žumanjak ima, kao što je gore spomenuto, značajan učinak na proces fragmentacije jajašca. Jajne stanice s malim udjelom žumanjka (oligolecital) potpuno se drobe, odnosno cjelokupna tvar oplođenog jajašca se tijekom drobljenja dijeli na nove stanice, blastomere (holoblastična jajašca). Naprotiv, u jajima koja sadrže više žumanjka ili čak veću količinu žumanjka (polilecital), brazde za cijepanje kontinuirano drobe samo manji dio ooplazme koji se nalazi na tzv. animalnom polu, gdje je manje žumanjčanih zrnaca (meroblastik). jaja).
U skladu s tim, pojedini predstavnici kategorije Kralježnjaci imaju sljedeće vrste drobljenja.
1. Potpuno drobljenje. Pod potpunim, totalnim drobljenjem podrazumijevaju se oni slučajevi kada se tijekom procesa drobljenja diobe cijela oplođena jajna stanica podijeli i brazde cijepanja se rašire po cijeloj njezinoj površini. Holoblastične jajne stanice drobe se ovom vrstom. Ovisno o sadržaju veće ili manje količine žumanjka u ooplazmi, kao i ovisno o njegovoj raspodjeli u ooplazmi, tijekom drobljenja pojavljuju se blastomeri ili relativno iste veličine (potpuno ravnomjerno, jednako ili adekvatno drobljenje), ili blastomeri različitih veličina, i to veće u području s visokim sadržajem žumanjka i manje na mjestu gdje je žumanjka manje (potpuno neravnomjerno, nejednako drobljenje). Veći blastomeri nazivaju se makromeri, a manji - mikromeri.
Puni jednaki, odn adekvatan, fragmentacija je karakteristična za oligolecitalna, izolecitalna jaja (lanceta, viših sisavaca i čovjeka); Mezolecitalne jajne stanice anizolecitalnih i umjereno telolecitalnih vrsta (neke niže ribe i vodozemci) fragmentirane su prema potpuno nejednakom tipu.
2. Djelomično, djelomično, drobljenje. Prema parcijalnom tipu, jajne stanice koje sadrže značajnu količinu žumanjka (polilecitalna jaja) su usitnjene, u kojima se zbog velike veličine Brazde cijepanja tijekom stanične diobe prodiru samo u područje životinjskog pola, gdje se nalazi stanična jezgra i gdje sloj ooplazme sadrži manje granula žumanjka (više ribe, gmazovi, ptice i neki niži sisavci, jajorodni).
S ovim drobljenje na animalnom polu relativno velikog jajeta zgnječeno je samo okruglo polje (disk), dok ostatak jajne stanice (kuglica žumanjka) ostaje nezgnječen (djelomično diskoidno cijepanje). Kod kukaca, iako su njihova polilecitalna centrolecitalna jaja zgnječena po cijeloj površini, središte stanice, koje sadrži veliku količinu žumanjka, ostaje nezgnječeno (djelomično površinsko drobljenje).
U navedenom crtanje prikazani su pojedini tipovi jajnih stanica ovisno o sadržaju i rasporedu žumanjka u ooplazmi, kao i ovisno o pripadajućem tipu cijepanja.
Proces kojim se zigota pretvara u višestanični organizam naziva se cijepanje. Ovo razdoblje je sljedeće nakon oplodnje i uključuje niz brojnih uzastopnih dioba.
Proces fragmentacije zigote traje oko šest dana. Sve stanice od kojih se sastoji embrij u medicini se nazivaju blastomeri. Fragmentacija zigote karakterizirana je vlastitim individualnim karakteristikama.
Interfaza, kao period, minimalnog je trajanja. Nakon toga slijede dvije potpune mitoze, što objašnjava progresivnu redukciju zigote. Do kraja šestog dana, nakon oplodnje, formirani višestanični organizam ne prelazi veličinu zigote. Ali proces fragmentacije završava u trenutku kada stanice embrija postanu slične somatskim stanicama ljudskog tijela.
Završni i Prva razina cijepanja zigota jedinstvena su po svojoj strukturi. U procesu kolosalnih promjena dolazi do punopravne asinkrone i podjednake podjele. Takvi podaci ukazuju na činjenicu da cijepanje zahvaća sve dijelove zigote, a blastomeri se pojavljuju iste veličine. Ako su stanice različite u volumenu, tada kada je zigota fragmentirana, dolazi do neistovremene mitotičke diobe.
Ne previše gust konglomerat stvaraju blastomeri otprilike u fazi razvoja zigote od osam stanica. Iako već šesti dan nakon oplodnje, otprilike nakon treće faze diobe, stanice stvaraju gustu strukturu unutar embrija. Taj se rad naziva zbijanjem i izaziva odvajanje unutarnjih blastomera od vanjskih. Vrste cijepanja zigote razlikuju se po razdoblju, a gore spomenuti stadij je morula. Takve središnje formacije stvaraju glavnu staničnu masu. A stanice povezane tijesnim spojevima služe kao neka vrsta barijere, koja je dizajnirana da zaštiti unutarnju strukturu morule. To jest, periferne stanice stvaraju trofoblast - vanjsku vrstu stanične mase.
Završni procesi fragmentacije zigote
Kao rezultat fragmentacije zigote, stanični organizam pretvara se u embrij. Tek četvrtog dana nakon oplodnje zigota ulazi u šupljinu maternice. U embriju se formira neka vrsta tekuće šupljine - blastocoel. Sada je embrij vezikula i ima ime - blastocista. Unutar tijela nalazi se stanični embrioblast - to je unutarnja masa. Iz te “materije” nastaje sam embrij i neki njegovi vanjski organi koji se vizualiziraju izvan embrija. Ako se unutarnja stanična masa počne dijeliti, tada će ta činjenica dovesti do stvaranja blizanaca.
Embrionalni dio posteljice nastaje na temelju trofoblasta koji stvara horion. Oko četvrtog dana nakon oplodnje stanice razaraju ovojnicu, odnosno mijenjaju prozirni dio embrija. Tako se zigota priprema za sljedeću fazu svoje transformacije.
Formira se zigota koja je sposobna za daljnji razvoj. Dioba zigote naziva se cijepanje. Razdvajanje- Ovo je ponovljena dioba zigote nakon oplodnje, uslijed čega nastaje višestanični embrij.
Zigota se vrlo brzo dijeli, stanice se smanjuju u veličini i nemaju vremena rasti. Stoga se embrij ne povećava u volumenu. Stanice koje nastaju kao rezultat nazivaju se blastomeri, a suženja koja ih odvajaju jedna od druge nazivaju se brazde cijepanja.
Na temelju smjera razlikuju se sljedeće brazde cijepanja: meridionalne – to su brazde koje dijele zigotu od animalnog do vegetativnog pola; ekvatorski žlijeb odvaja zigotu duž ekvatora; širinski žlijebovi idu paralelno s ekvatorijalnim žlijebom; tangencijalni žljebovi idu paralelno s površinom zigote.
Uvijek postoji jedan ekvatorijalni žlijeb, ali može biti više meridionalnih, latitudinalnih i tangencijalnih žlijebova. Smjer cijepnih brazda uvijek je određen položajem cijepnog vretena.
Drobljenje se uvijek odvija prema određenim pravilima:
Prvo pravilo odražava položaj vretena cijepanja u blastomeri, naime:
– vreteno cijepanja nalazi se prema najvećem dijelu citoplazme, bez inkluzija.
Drugo pravilo odražava smjer drobljenja brazda:
– brazde cijepanja uvijek idu okomito na vreteno cijepanja.
Treće pravilo odražava brzinu prolaska brazda za drobljenje:
– brzina prolaska brazdi cijepanja obrnuto je proporcionalna količini žumanjka u jajetu, tj. u onom dijelu stanice gdje ima malo žumanjka, brazde će prolaziti većom brzinom, a u dijelu gdje ima više žumanjka, brzina prolaska cijepnih brazda će se usporiti.
Cijepanje ovisi o količini i položaju žumanjka u jajetu. S malom količinom žumanjka zgnječi se cijela zigota, s većom količinom zgnječi se samo dio zigote bez žumanjka. S tim u vezi jaja se dijele na holoblastična (potpuno zgnječena) i meroblastna (djelomično zgnječena). Slijedom toga, drobljenje ovisi o količini žumanjka i, uzimajući u obzir niz karakteristika, dijeli se: prema potpunosti procesa pokrivanja materijala zigote na potpuno i nepotpuno; u odnosu na veličinu nastalih blastomera na jednolike i neravnomjerne te u pogledu konzistencije dioba blastomera - sinkrone i asinkrone.
Potpuno drobljenje može biti ravnomjerno ili neravnomjerno. Potpuno ujednačeno svojstvo je jaja s malom količinom žumanjka i njegovim više-manje ujednačenim smještajem u žumanjku. Ova vrsta cijepanja jajeta. U tom slučaju ide prva brazda od animalnog do vegetativnog pola, a nastaju dva blastomera; drugi žlijeb je također meridionalni, ali ide okomito na prvi, i formiraju se četiri blastomera. Treći je ekvatorijalni, formira se osam blastomera. Nakon toga dolazi do izmjene meridionalnih i latitudinalnih brazda drobljenja. Broj blastomera nakon svake diobe povećava se dva puta (2; 4; 16; 32 itd.). Kao rezultat takve fragmentacije nastaje sferni embrij, koji se naziva blastula. Stanice koje tvore stijenku blastule nazivaju se blastoderm, a unutarnja šupljina naziva se blastocel. Životinjski dio blastule naziva se krov, a vegetativni dio dno blastule.

Potpuna neravnomjerna fragmentacija tipična je za jaja s prosječnim sadržajem žumanjka koja se nalaze u vegetativnom dijelu. Takva su jaja karakteristična za ciklostome i. pri čemu vrsta drobljenja nastaju blastomeri nejednake veličine. U animalnom polu nastaju male blastomere koje nazivamo mikromere, a u vegetativnom polu velike blastomere – makromere. Prve dvije brazde, kao i one kod lanceleta, idu meridionalno; treći žlijeb odgovara ekvatorijalnom žlijebu, ali je pomaknut od ekvatora prema životinjskom polu. Budući da animalni pol sadrži citoplazmu bez žumanjka, ovdje dolazi do brže fragmentacije i stvaranja malih blastomera. Vegetativni pol sadrži glavninu žumanjka, pa brazde cijepanja prolaze sporije i stvaraju se veliki blastomeri.
Nepotpuno cijepanje je karakteristično za telolecitalna i centrolecitalna jaja. U drobljenju sudjeluje samo onaj dio jajeta koji je slobodan od žumanjka. Nepotpuno drobljenje dijelimo na diskoidno (teleosti, gmazovi, ptice) i površinsko (člankonošci).
Telolecitalna jaja, kod kojih je velika količina žumanjka koncentrirana u vegetativnom dijelu, podijeljena su nepotpunim diskoidnim cijepanjem. U tim je jajima dio citoplazme bez žumanjka u obliku klicinog diska raširen na žumanjku na animalnom polu. Cijepanje se događa samo u području zametnog diska. Vegetativni dio jajeta, ispunjen žumanjkom, ne sudjeluje u drobljenju. Debljina zametnog diska je beznačajna, stoga su cijepna vretena tijekom prva četiri dijeljenja smještena vodoravno, a cijepne brazde idu okomito. Formira se jedan red stanica. Nakon nekoliko dioba stanice postaju visoke i cijepna vretena se u njima nalaze okomito, a cijepne brazde idu paralelno s površinom jajeta. Kao rezultat toga, klica se pretvara u ploču koja se sastoji od nekoliko redova stanica. Između zametnog diska i žumanjka pojavljuje se mala šupljina u obliku proreza, koja je slična blastocelu.
Nepotpuno površinsko cijepanje uočeno je u centrolecitalnim jajima s velikom količinom žumanjka u sredini. Citoplazma u takvim jajima nalazi se duž periferije, a manji dio u središtu blizu jezgre. Ostatak ćelije napunimo žumanjkom. Kroz masu žumanjka prolaze tanke citoplazmatske niti koje povezuju perifernu citoplazmu s perinuklearnom. Fragmentacija počinje fisijom jezgri, što rezultira povećanjem broja jezgri. Okruženi su tankim rubom citoplazme, pomiču se prema periferiji i nalaze se u citoplazmi bez žumanjka. Čim jezgre uđu u površinski sloj, on se prema njihovom broju dijeli na blastomere. Kao rezultat takve fragmentacije, cijeli središnji dio citoplazme pomiče se na površinu i stapa se s perifernim dijelom. Izvana se formira čvrsti blastoderm iz kojeg se razvija embrij, a unutra se nalazi žumanjak. Površinska fragmentacija karakteristična je za jaja člankonožaca.
Na prirodu fragmentacije utječu i svojstva citoplazme, koja određuju relativni položaj blastomera. Na temelju ove značajke razlikuju se radijalno, spiralno i bilateralno drobljenje. Kod radijalnog drobljenja, svaki gornji blastomer nalazi se točno ispod donjeg (koelenterati, bodljikaši, lancelet itd.). Tijekom spiralnog drobljenja, svaki gornji blastomer pomaknut je u odnosu na donji za pola, tj. svaki gornji blastomer nalazi se između dva donja. U ovom slučaju, blastomeri su raspoređeni kao u spirali (crvi, mekušci). S bilateralnim drobljenjem, kroz zigotu se može povući samo jedna ravnina, s obje strane koje će se uočiti identični blastomeri (valjkasti crvi, ascidije).