Začiatok nového organizmu je daný oplodneným vajíčkom (s výnimkou prípadov partenogenézy a vegetatívneho rozmnožovania). Hnojenie je proces vzájomného splynutia dvoch zárodočných buniek (gamét), počas ktorého sa vykonávajú dve rôzne funkcie: sexuálna (spojenie génov dvoch rodičovských jedincov) a reprodukčná (vznik nového organizmu). Prvá z týchto funkcií zahŕňa prenos génov z rodičov na potomkov, druhá - spustenie tých reakcií a pohybov v cytoplazme vajíčka, ktoré umožňujú ďalší vývoj. V dôsledku oplodnenia sa vo vajíčku obnoví dvojitá (2p) sada chromozómov. Spermiou zavedený centrozóm po zdvojení tvorí štiepne vreteno a zygota vstupuje do 1. štádia embryogenézy – štádia drvenia. V dôsledku mitózy sa zo zygoty vytvoria 2 dcérske bunky - blastoméry.
Prezygotické obdobie
Prezygotické obdobie vývoja je spojené s tvorbou gamét (gametogenéza). Tvorba oocytov začína u žien ešte pred narodením a je dokončená pre každý daný oocyt až po jeho oplodnení. Ženský plod vo vaječníkoch obsahuje v čase narodenia asi dva milióny oocytov prvého rádu (sú to ešte diploidné bunky) a len 350 - 450 z nich dosiahne štádium oocytov druhého rádu (haploidné bunky), pričom do vajec (jedno po druhom počas jednej menštruačný cyklus). Na rozdiel od žien sa zárodočné bunky v semenníkoch (semenníkoch) u mužov začínajú vytvárať až s nástupom puberty. Trvanie obdobia tvorby spermií je približne 70 dní; na jeden gram hmotnosti semenníkov je počet spermií asi 100 miliónov denne.
Hnojenie
Hnojenie - splynutie samčej pohlavnej bunky (spermie) so samicou (vajíčko, vajíčko), čo vedie k vytvoreniu zygoty - nového jednobunkového organizmu. Biologickým významom oplodnenia je zjednotenie jadrového materiálu mužských a ženských gamét, čo vedie k zjednoteniu otcovských a materských génov, obnove diploidnej sady chromozómov, ako aj k aktivácii vajíčka, tj. jeho stimulácia pre embryonálny vývoj. Spojenie vajíčka so spermiou sa zvyčajne vyskytuje v lievikovitej časti vajíčkovodu počas prvých 12 hodín po ovulácii.
Semenná tekutina, vstupujúca do pošvy ženy pri pohlavnom styku, obsahuje zvyčajne 60 až 150 miliónov spermií, ktoré vďaka pohybom rýchlosťou 2-3 mm za minútu, neustálym zvlneným kontrakciám maternice a trubíc a zásaditému prostrediu, už po 1-2 minútach po pohlavnom styku sa dostanú do maternice a po 2-3 hodinách do koncových úsekov vajíčkovodov, kde sa zvyčajne spájajú s vajíčkom. Existuje monospermické (jedna spermia vstúpi do vajíčka) a polyspermické (dve alebo viac spermií vstúpi do vajíčka, ale iba jedno jadro spermie sa spojí s jadrom vajíčka). Zachovanie aktivity spermií pri ich prechode v pohlavnom trakte ženy uľahčuje mierne zásadité prostredie krčka maternice vyplnené hlienovou zátkou. Pri orgazme pri pohlavnom styku sa slizničná zátka z krčka maternice čiastočne vytlačí a následne sa do nej stiahne a tým prispieva k rýchlejšiemu vstupu spermií z pošvy (kde je normálne napr. zdravá žena mierne kyslé prostredie) do priaznivejšieho prostredia krčka maternice a dutiny maternice. Prechod spermií cez hlienovú zátku cervikálneho kanála je tiež uľahčený prudkým zvýšením priepustnosti hlienu v dňoch ovulácie. Vo zvyšných dňoch menštruačného cyklu má hlienová zátka výrazne nižšiu priepustnosť pre spermie.
Mnohé spermie nachádzajúce sa v pohlavnom trakte ženy si môžu zachovať schopnosť oplodnenia 48-72 hodín (niekedy až 4-5 dní). Ovulované vajíčko zostáva životaschopné približne 24 hodín. Vzhľadom na to je najpriaznivejším časom na oplodnenie obdobie prasknutia zrelého folikulu, po ktorom nasleduje narodenie vajíčka, ako aj 2-3 deň po ovulácii. Ženy používajúce fyziologickú metódu antikoncepcie by si mali uvedomiť, že načasovanie ovulácie môže kolísať a životaschopnosť vajíčka a spermie môže byť výrazne dlhšia. Krátko po oplodnení sa začína štiepenie zygoty a tvorba embrya.
ZygotaZygota (grécky párová zygota) je diploidná (obsahujúca kompletnú dvojitú sadu chromozómov) bunka, ktorá je výsledkom oplodnenia (fúzie vajíčka a spermie). Zygota je totipotentná (to znamená schopná produkovať akúkoľvek inú) bunku. Termín zaviedol nemecký botanik E. Strasburger.
U ľudí nastáva prvé mitotické delenie zygoty približne 30 hodín po oplodnení, čo je spôsobené zložitými procesmi prípravy na prvý akt drvenia. Bunky vytvorené v dôsledku drvenia zygoty sa nazývajú blastoméry. Prvé delenia zygoty sa nazývajú „drvenie“, pretože bunka je rozdrvená: po každom delení sa dcérske bunky zmenšujú a medzi deleniami neexistuje štádium bunkového rastu.
Vývoj zygoty Zygota sa buď začína vyvíjať ihneď po oplodnení, alebo je oblečená v hustej škrupine a na nejaký čas sa zmení na pokojnú spóru (často nazývanú zygospóra) - typickú pre mnohé huby a riasy.
RozdelenieObdobie embryonálny vývoj mnohobunkový živočích začína rozdrvením zygoty a končí narodením nového jedinca. Proces štiepenia pozostáva zo série postupných mitotických delení zygoty. Dve bunky vytvorené v dôsledku nového delenia zygoty a všetky nasledujúce generácie buniek v tomto štádiu sa nazývajú blastoméry. Pri drvení nasleduje jedno delenie za druhým a vzniknuté blastoméry nerastú, v dôsledku čoho je každá nová generácia blastomér reprezentovaná menšími bunkami. Táto vlastnosť bunkového delenia počas vývoja oplodneného vajíčka určila vzhľad obrazového pojmu - rozdrvenie zygoty.
O odlišné typyŽivočíšne vajcia sa líšia množstvom a charakterom distribúcie rezervných živín (žĺtka) v cytoplazme. To do značnej miery určuje povahu následnej fragmentácie zygoty. Pri malom množstve a rovnomernej distribúcii žĺtka v cytoplazme sa celá hmota zygoty rozdelí za vzniku identických blastomérov - úplné rovnomerné rozdrvenie (napríklad u cicavcov). Keď sa žĺtok hromadí prevažne na jednom z pólov zygoty, dochádza k nerovnomernej fragmentácii – vznikajú blastoméry, ktoré sa líšia veľkosťou: väčšie makroméry a mikroméry (napríklad u obojživelníkov). Ak je vajíčko veľmi bohaté na žĺtok, potom sa jeho časť bez žĺtka rozdrví. Takže u plazov, vtákov, iba diskovitá časť zygoty na jednom z pólov, kde sa nachádza jadro, podlieha drveniu - neúplnému, diskoidnému drveniu. Nakoniec u hmyzu sa do procesu drvenia zúčastňuje iba povrchová vrstva cytoplazmy zygoty - neúplné, povrchové drvenie.
V dôsledku drvenia (keď počet deliacich sa blastomérov dosiahne významný počet) sa vytvorí blastula. V typickom prípade (napríklad v lancelete) je blastula dutá guľa, ktorej stena je tvorená jednou vrstvou buniek (blastoderm). Dutina blastuly – blastocoel, inak nazývaná primárna telesná dutina, je naplnená tekutinou. U obojživelníkov má blastula veľmi malú dutinu a u niektorých živočíchov (napríklad článkonožcov) môže blastocoel úplne chýbať.
gastruláciaV ďalšom štádiu embryonálneho obdobia prebieha proces tvorby gastruly - gastrulácia. U mnohých zvierat dochádza k tvorbe gastruly invagináciou, t.j. výbežky blastodermu na jednom z pólov blastuly (s intenzívnou reprodukciou buniek v tejto zóne). V dôsledku toho sa vytvorí dvojvrstvové embryo v tvare misky. Vonkajšia vrstva buniek je ektoderm a vnútorná vrstva je endoderm. Vnútorná dutina, ktorá vzniká pri vyčnievaní steny blastuly, primárne črevo, komunikuje s vonkajším prostredím cez otvor – primárne ústa (blastopor). Existujú aj iné typy gastrulácie. Napríklad u niektorých coelenterátov sa endoderm gastruly tvorí imigráciou, t.j. „vypudenie“ časti blastodermálnych buniek do dutiny blastuly a ich následné rozmnoženie. Primárne ústie vzniká pretrhnutím steny gastruly. Pri nerovnomernom drvení (u niektorých červov, mäkkýšov) sa gastrula vytvára v dôsledku zanášania makromérov mikromérmi a tvorby endodermy v dôsledku prvej. Často sa kombinujú rôzne metódy gastrulácie.
U všetkých živočíchov (okrem húb a koelenterátov - dvojvrstvových živočíchov) končí štádium gastrulácie vytvorením ďalšej vrstvy buniek - mezodermu. Táto "bunková vrstva sa tvorí medzi ento- a ektodermou. Sú známe dva spôsoby kladenia mezodermy. Napríklad u annelidov sú v oblasti blastopóru gastruly izolované dve veľké bunky (teloblasty). Rozmnožujú sa a dávajú vznik na dva mezodermálne pruhy, z ktorých (čiastočne v dôsledku divergencie buniek, čiastočne v dôsledku deštrukcie časti buniek vo vnútri mezodermálnych pruhov) vznikajú coelomické vaky - teloblastický spôsob kladenia mezodermu.Pri enterocelóznej metóde (ostnatokožce, lanceleta, stavovce) sa v dôsledku vyčnievania steny primárneho čreva vytvoria bočné vrecká, ktoré sa následne oddelia a stanú sa coelomickými. mezodermálna vrstva buniek vystielajúca vnútro telovej dutiny tvorí peritoneálny epitel Dutina, ktorá tak nahradila primárnu, sa nazýva sekundárna telová dutina alebo coelom Pri teloblastickej metóde iniciácie mezodermu sa blastopór vyvíja do ústneho otvoru dospelého zvieraťa. Takéto organizmy sa nazývajú protostómy. V deuterostómoch (s enterocoelóznou metódou kladenia mezodermu) blastopór prerastá alebo sa mení na konečník a ústa dospelého človeka sa vyskytujú druhýkrát, vyčnievaním ektodermy.
Tvorba troch zárodočných vrstiev (ekto-, ento- a mezoderm) završuje štádium gastrulácie a od tohto momentu začínajú procesy histo- a organogenézy. V dôsledku bunkovej diferenciácie troch zárodočných vrstiev vznikajú rôzne tkanivá a orgány vyvíjajúceho sa organizmu. Už koncom minulého storočia (najmä vďaka štúdiám I. I. Mečnikova a A. O. Kovalevského) sa zistilo, že u rôznych živočíšnych druhov z rovnakých zárodočných vrstiev vznikajú rovnaké orgány a tkanivá. Z ektodermy sa tvorí epidermis so všetkými odvodenými štruktúrami a nervový systém. Vďaka endodermu sa tvorí tráviaci trakt a pridružené orgány (pečeň, pankreas, pľúca atď.). Mezoderm tvorí kostru, cievny systém, vylučovací aparát, pohlavné žľazy. Zárodočné vrstvy sa dnes síce nepovažujú za prísne špecializované, no napriek tomu je u prevažnej väčšiny živočíšnych druhov zjavná ich homológia, čo svedčí o jednote pôvodu živočíšnej ríše.
Počas embryonálneho obdobia dochádza u vyvíjajúcich sa organizmov k zvýšeniu rýchlosti rastu a diferenciácie. Ak počas štiepenia nedôjde k rastu a blastula (z hľadiska hmotnosti) môže byť výrazne nižšia ako zygota, potom, počnúc procesom gastrulácie, sa hmotnosť embrya rýchlo zvyšuje (v dôsledku intenzívnej reprodukcie buniek). Procesy bunkovej diferenciácie začínajú v najskoršom štádiu embryogenézy - drvenie a sú základom primárnej diferenciácie tkanív - vznik troch zárodočných vrstiev (embryonálnych tkanív). Ďalší vývoj embrya sprevádza stále sa zväčšujúci proces diferenciácie tkanív a orgánov. V dôsledku embryonálneho obdobia vývoja vzniká organizmus, ktorý je schopný samostatnej (viac či menej) existencie vo vonkajšom prostredí. Nový jedinec sa narodí buď ako výsledok vyliahnutia z vajíčka (u vajcorodých zvierat) alebo výstupom z tela matky (u viviparóz).
Histo - a organogenézaHisto- a organogenéza embrya sa uskutočňuje v dôsledku reprodukcie, migrácie, diferenciácie buniek, ich zložiek, vytvárania medzibunkových kontaktov a smrti niektorých buniek. 317. až 20. deň pokračuje presomitickým obdobím od 20. dňa začína obdobie vývoja somitu. Na 20. deň embryogenézy sa prostredníctvom tvorby kmeňových záhybov (cefalokaudálnych a laterálnych) oddelí samotné embryo od extraembryonálnych orgánov, ako aj jeho plochý tvar sa zmení na valcovitý. Zároveň sú dorzálne časti mezodermu embrya rozdelené na samostatné segmenty umiestnené na oboch stranách chordy - somity. Na 21. deň sú v tele embrya 2-3 páry somitov. Somity sa začínajú vytvárať z páru III, páry I a II sa objavujú o niečo neskôr. Počet somitov sa postupne zvyšuje: na 23. deň vývinu je 10 párov somitov, 25. - 14 párov, 27. - 25 párov, na konci piateho týždňa dosahuje počet somitov v zárodku 43. - 44 párov. Na základe výpočtu počtu somitov je možné približne určiť načasovanie vývoja (somitský vek) embrya.
Z vonkajšej časti každého somitu vzniká dermatóm, z vnútornej - sklerot, zo stredu - myot. Dermatóm sa stáva zdrojom kožnej dermy, sklerotóm sa stáva zdrojom chrupavkového a kostného tkaniva a myotóm sa stáva zdrojom kostrových svalov dorzálnej časti embrya. Ventrálne časti mezodermu - splanchnotóm - nie sú segmentované, ale sú rozdelené na viscerálne a parietálne listy, z ktorých sa vyvíjajú serózne membrány vnútorných orgánov, sval srdca a kôry nadobličiek. Z mezenchýmu splanchnotómu sa tvoria krvné cievy, krvné bunky, spojivové a hladké svalové tkanivo embrya. Úsek mezodermu, ktorý spája somity so splanchnotómom, je rozdelený na segmentované nohy - nefrogonot, ktoré slúžia ako zdroj pre vývoj obličiek a pohlavných žliaz, ako aj paramezonefrických vývodov. Z posledného sa tvorí epitel maternice a vajcovodu.
V procese diferenciácie zárodočnej ektodermy vzniká neurálna trubica, neurálne hrebene, plakody, kožná ektoderma a prechordálna platnička. Proces tvorby neurálnej trubice sa nazýva neurulácia. Spočíva vo vytvorení štrbinovitej priehlbiny na povrchu ektodermy; zhrubnuté okraje tejto priehlbiny (neurálne záhyby) sa spájajú a vytvárajú nervovú trubicu. Mozgové vezikuly sa tvoria z lebečnej časti nervovej trubice, ktorá je základom mozgu. Na oboch stranách nervovej trubice (medzi poslednou a kožným ektodermom) sú oddelené skupiny buniek, z ktorých sa vytvárajú nervové hrebene. Bunky neurálnej lišty sú schopné migrovať. Z buniek migrujúcich v smere dermatómu vznikajú pigmentové bunky - melanocyty; bunky neurálnej lišty, ktoré migrujú smerom k brušnej dutine, vedú k vzniku sympatických a parasympatických ganglií, drene nadobličiek. Z buniek neurálnych hrebeňov, ktoré nemigrovali, vznikajú gangliové platničky, z ktorých sa vyvíjajú spinálne a periférne autonómne nervové gangliá. Z plakov sa tvoria gangliá hlavy a nervové bunky orgánu sluchu a rovnováhy.
Rozdelenie(segmentácia) u jednotlivých zástupcov kategórie stavovcov má spravidla rovnaký priebeh; ako však už bolo spomenuté vyššie, ovplyvňovali ho faktory, ktoré počas fylogenézy ovplyvňovali vývoj v podobe dôsledkov vplyvu vnútorného a vonkajšieho prostredia, v ktorom organizmy žili počas svojho rodového vývoja (cenogenetické faktory).
Pri pozorovaní zmeny, vyskytujúcich sa vo vajciach podľa fylogenetického vývoja vajíčok jednotlivých zástupcov kategórie stavovcov, je vidieť, že vaječné bunky sa od seba výrazne líšia obsahom živnej a stavebnej látky - žĺtka. Vaječné bunky kopinatca (Amfioxus), organizmu, ktorý je fylogeneticky považovaný za najnižšie organizovaného tvora, ktorý však už má silnú chrbtovú oblasť, patria medzi oligoleciálne.
Avšak v súlade s fylogenetickým vývojom Množstvo žĺtka vo vajciach stavovcov, ktoré sú fylogeneticky najorganizovanejšími organizmami, sa stále viac a viac zvyšuje, pričom maximálne množstvo dosahuje vo vajciach vtákov, ktoré sú relatívne veľmi veľké a polylecitálne. Vplyvom koenogenetických faktorov (faktory ovplyvňujúce z vonkajšieho prostredia a determinované zmenami životného štýlu a následne aj vývoja) sa množstvo žĺtka v procese fylogenetického vývoja smerom k človeku stále viac znižuje, čím sa vajíčka ľudí a vyššie cicavce sa stávajú opäť (sekundárne) oligolecitálnymi .
S premenlivou sumou žĺtok má, ako bolo uvedené vyššie, významný vplyv na proces drvenia vajíčka. Vaječné bunky s nízkym obsahom žĺtka (oligolecitálne) sú úplne rozdrvené, to znamená, že celá látka oplodneného vajíčka sa pri drvení rozdelí na nové bunky, blastoméry (vajíčka holoblastického typu). Naopak, vo vajciach obsahujúcich viac žĺtka, alebo dokonca veľké množstvo žĺtka (polylecitálne), štiepne brázdy priebežne drvia len menšiu časť ooplazmy, ktorá sa nachádza na takzvanom zvieracom póle, kde je menej žĺtkových granúl (meroblastické typ vajec).
V súlade s týmto jednotlivých zástupcov kategórie stavovce, rozlišujú sa tieto druhy drvenia.
1. Úplné rozdrvenie. Úplné, úplné rozdrvenie zahŕňa tie prípady, keď sa v procese rozdrvenia delí celá oplodnená vaječná bunka a po celom jej povrchu sa rozprestierajú drvené ryhy. Podľa tohto typu sú vaječné bunky holoblastických druhov rozdrvené. V závislosti od obsahu väčšieho alebo menšieho množstva žĺtka v ooplazme, ako aj v závislosti od jeho distribúcie v ooplazme sa pri drvení môžu vyskytovať blastoméry buď relatívne rovnakej veľkosti (úplne rovnomerné, rovnaké alebo rovnaké štiepenie) alebo blastoméry zn. rôzne veľkosti, a to väčšie v oblasti s vysokým obsahom žĺtka a menšie v mieste, kde je žĺtka menej (úplné nerovnomerné, nerovnomerné drvenie). Väčšie blastoméry sa nazývajú makroméry, menšie sa nazývajú mikroméry.
Úplne rovné, príp primeraný, drvenie je charakteristické pre vajíčka oligolecitálne, izolecitálne (lancelet, vyššie cicavce a ľudia); podľa úplne nerovnomerného typu sú rozdrvené mesolecitálne vaječné bunky anisolecitálneho a stredne telolecitného druhu (niektoré nižšie ryby a obojživelníky).
2. Čiastočné, čiastočné, zdrvujúce. Čiastočným typom sú vaječné bunky rozdrvené, obsahujúce značné množstvo žĺtka (polylecitálne vajcia), ktoré v dôsledku ich veľké veľkostištiepne ryhy pri delení buniek prenikajú len do oblasti zvieracieho pólu, kde sa nachádza bunkové jadro a kde vrstva ooplazmy obsahuje menej žĺtkových granúl (vyššie ryby, plazy, vtáky a niektoré nižšie cicavce, vajcorodé).
S takými drvenie na zvieracom póle pomerne veľkého vajca sa rozdrví iba okrúhle pole (disk), zatiaľ čo zvyšok vaječnej bunky (žĺtková guľa) zostane nerozbitý (čiastočné diskovité drvenie). U hmyzu ich polyleciálne centrolecitálne oocyty sú síce rozdrvené po celom povrchu, ale stred bunky, obsahujúci veľké množstvo žĺtka, zostáva nerozdrvený (čiastočné povrchové drvenie).
Vo vyššie uvedenom obrázok jednotlivé typy vaječných buniek sú zobrazené v závislosti od obsahu a rozloženia žĺtka v ooplazme, ako aj v závislosti od zodpovedajúceho typu drvenia.
Embryogenéza (grécky zárodok - embryo, genéza - vývoj) - rané obdobie individuálny rozvoj organizmu od okamihu oplodnenia (počatia) až po pôrod, je počiatočná fáza ontogenéza (gr. ontos - bytie, genesis - vývoj), proces individuálneho vývoja organizmu od počatia až po smrť.
Vývoj akéhokoľvek organizmu začína ako výsledok fúzie dvoch pohlavných buniek (gamét), mužskej a ženskej. Všetky bunky tela, napriek rozdielom v štruktúre a funkciách, spája jedno – jediná genetická informácia uložená v jadre každej bunky, jedna dvojitá sada chromozómov (okrem vysoko špecializovaných krviniek – erytrocytov, ktoré nemajú majú jadro). To znamená, že všetky somatické (soma - telové) bunky sú diploidné a obsahujú dvojitú sadu chromozómov - 2 n, a iba zárodočné bunky (gaméty), ktoré sa tvoria v špecializovaných pohlavných žľazách (semenníky a vaječníky), obsahujú jednu sadu chromozómov - 1 n.
Keď sa pohlavné bunky spoja, vytvorí sa bunka - zygota, v ktorej sa obnoví dvojitá sada chromozómov. Pripomeňme, že jadro ľudskej bunky obsahuje 46 chromozómov, respektíve zárodočné bunky majú 23 chromozómov.
Pohlavie dieťaťa je určené pomerom chromozómov počas oplodnenia. Ak je vajíčko oplodnené spermiou s pohlavným chromozómom X, potom sa v zygote vytvoria dva chromozómy x ( ženské telo). Pri oplodnení spermie s chromozómom y sa v zygote vytvorí kombinácia XY (mužské telo). Keďže meióza produkuje rovnaký počet spermií s chromozómami X a Y, teoreticky by mal byť počet novonarodených chlapcov a dievčat rovnaký, ale v skutočnosti sa na 100 chlapcov narodí 103 dievčat. Je to spôsobené väčšou citlivosťou plodov mužského pohlavia na rôzne nepriaznivé a škodlivé faktory.
Typy embryogenézy

Typ embryogenézy je súbor znakov, ktoré poskytujú vyvíjajúcemu sa organizmu spojenie s prostredím.
nelarválneho typu Veľké vajcia, veľa žĺtkov. Embryá sú dlhodobo chránené vaječnými membránami s využitím zásob živín uložených vo vajíčku. Žraloky, raje, škrkavky a ploché červy, množstvo hmyzu a plazov, vtáky a vajcorodé cicavce.
Sekundárny typ lariev- vajíčka sú malé, z vajíčok vychádzajú pohyblivé embryá, schopné kŕmenia. Vývoj je chránený špeciálne vzdelanie(kapsuly), v prípade živorodenia - v tele matky. Živé narodenie je charakteristické pre placentu, tropické škorpióny, vačnaté cicavce a niektoré ryby a hmyz.
Periodizácia ontogenézy: zygota, štiepenie, gastrulácia, histogenéza a organogenéza
Zygota

Zygota, ktorá vznikla ako výsledok fúzie ženských a mužských gamét, je jednobunkovým štádiom vývoja mnohobunkového organizmu.
V zygote je možné vysledovať výrazné pohyby cytoplazmy, v dôsledku čoho sa určujú oblasti, z ktorých sa v budúcnosti vyvinú určité orgány a tkanivá.
Stanovujú sa v ňom samostatné úseky cytoplazmy, syntetizuje sa DNA a proteíny. Zygota má bisymetrickú štruktúru. Postupne dochádza k porušeniu pomeru jadra a cytoplazmy, v dôsledku čoho sa stimuluje proces delenia - drvenia
Štiepenie + blastula
Drvenie vykonáva nasledujúce funkcie:
- vzniká dostatočný počet buniek potrebných na tvorbu tkanív a orgánov.
- redistribúcia žĺtka a cytoplazmy medzi dcérskymi bunkami. 1 a 2 deliace brázdy vedú pozdĺž poludníka a 3 pozdĺž rovníka. Bližšie k zvieraciemu pólu.
- určuje sa plán embrya - dorzálno-brušná os, predo-zadná os.
- jadrovo-cytoplazmatické vzťahy sú normalizované. Počet jadier rastie, objem a hmotnosť zostávajú rovnaké.
Vlastnosti drvenia:
- medzifázy sú krátke
- blastoméry nerastú
- protoplazma sa delí štiepnymi brázdami
Rozdelenie – ide o sériu postupných mitotických delení zygoty a ďalších blastomér, ktoré končia vytvorením mnohobunkového embrya - blastula .
Prvé rozdelenie štiepenia začína po spojení dedičného materiálu pronuklea a vytvorení spoločnej metafázovej platničky. Bunky vytvorené počas štiepenia sa nazývajú blastoméry. Znakom mitotických delení drvenia je, že s každým delením sa bunky zmenšujú a zmenšujú, až kým nedosiahnu pomer objemov jadra a cytoplazmy, ktorý je obvyklý pre somatické bunky. O morský ježko, napríklad vyžaduje šesť delení a embryo pozostáva zo 64 buniek. Medzi po sebe nasledujúcimi deleniami nedochádza k rastu buniek, ale DNA sa nevyhnutne syntetizuje.
Všetky prekurzory DNA a potrebné enzýmy sa akumulujú počas oogenézy. V dôsledku toho sa mitotické cykly skracujú a delenia nasledujú po sebe oveľa rýchlejšie ako v bežných somatických bunkách. Po prvé, blastoméry sú priľahlé k sebe a tvoria zhluk buniek nazývaný morula. Potom sa medzi bunkami vytvorí dutina - blastocoel, naplnená tekutinou. Bunky sú vytlačené na perifériu, čím sa vytvorí stena blastuly - blastoderm. Celková veľkosť embrya na konci štiepenia v štádiu blastuly nepresahuje veľkosť zygoty.
Hlavným výsledkom zdrvujúceho obdobia je premena zygoty na mnohobunkové jednozmenné embryo.
Morfológia drvenia
Spravidla sú blastoméry usporiadané v prísnom poradí voči sebe navzájom a voči polárnej osi vajíčka. Poradie alebo spôsob drvenia závisí od množstva, hustoty a rozloženia žĺtka vo vajci. Podľa Sachs-Hertwigových pravidiel má bunkové jadro tendenciu byť umiestnené v strede cytoplazmy bez žĺtka a vreteno bunkového delenia má tendenciu byť v smere najväčšieho rozsahu tejto zóny.
V oligo- (malý žĺtok) a mesolecitálnych (priemerné množstvo žĺtka) vajciach drvenie kompletný, alebo holoblastický. Tento typ drvenia sa vyskytuje u mihule, niektorých rýb, všetkých obojživelníkov, ako aj u vačkovcov a placentárnych cicavcov. Pri úplnom rozdrvení zodpovedá rovina prvého delenia rovine obojstrannej symetrie. Rovina druhého delenia prebieha kolmo na rovinu prvého. Obe brázdy prvých dvoch divízií sú poludníkové, t.j. začínajú na zvieracom póle a šíria sa k vegetatívnemu pólu. Vaječná bunka je rozdelená na štyri blastoméry, ktoré majú viac-menej rovnakú veľkosť. Rovina tretieho delenia prebieha kolmo na prvé dve v smere zemepisnej šírky. Potom sa v mezolecitálnych vajíčkach v štádiu ôsmich blastomérov prejavuje nerovnomerné drvenie. Na živočíšnom póle sú štyri menšie blastoméry - mikroméry, na vegetatívnom póle - štyri väčšie - makroméry. Potom rozdelenie opäť ide v rovinách poludníka a potom znova v rovinách zemepisnej šírky.
V polylecitálnych (veľa žĺtkov) vajciach kostnatých rýb, plazov, vtákov a tiež monotrémnych cicavcov je fragmentácia čiastočná, alebo meroblastická, t.j. pokrýva iba cytoplazmu bez žĺtka. Nachádza sa vo forme tenkého disku pri zvieracom póle, preto sa tento typ drvenia nazýva diskoidálny.
Pri charakterizácii typu drvenia sa berie do úvahy aj relatívna poloha a rýchlosť delenia blastomérov. Ak sú blastoméry usporiadané v radoch nad sebou pozdĺž polomerov, drvenie sa nazýva radiálne. Je typický pre strunatce a ostnatokožce. V prírode existujú iné varianty priestorového usporiadania blastomérov počas drvenia, ktoré určuje také typy, ako sú špirálové u mäkkýšov, bilaterálne u ascaris, anarchické u medúz.
Na konci drvenia, blastula. Typ blastuly závisí od typu drvenia, a teda od typu vajíčka.
gastrulácia

V procese gastrulácie možno rozlíšiť 2 fázy:
- tvorba ekto- a endodermu (2-vrstvové embryo)
- tvorba mezodermu (3-vrstvové embryo)
Podstatou štádia gastrulácie je, že jednovrstvové embryo – blastula – sa mení na viacvrstvové – dvoj- alebo trojvrstvové, nazývané gastrula.
U primitívnych strunatcov, napríklad v lancelete, sa homogénna jednovrstvová blastoderm počas gastrulácie premení na vonkajšiu zárodočnú vrstvu - ektodermu - a vnútornú zárodočnú vrstvu - endodermu.
Endoderm tvorí primárne črevo s dutinou vo vnútri gastrocoelu. Otvor vedúci k gastrocoelu sa nazýva blastopór alebo primárne ústa. Rozhodujúce sú dve zárodočné vrstvy morfologické znaky gastrulácia. Ich existencia v určitom štádiu vývoja u všetkých mnohobunkových živočíchov, od koelenterátov až po vyššie stavovce, nám umožňuje uvažovať o homológii zárodočných vrstiev a jednote pôvodu všetkých týchto živočíchov. U stavovcov sa pri gastrulácii okrem spomínaných dvoch tvorí aj tretia zárodočná vrstva - mezoderm, ktorá zaberá miesto medzi ekto- a endodermou.
Vývoj strednej zárodočnej vrstvy, ktorou je chordomesoderm, je evolučnou komplikáciou gastrulačnej fázy u stavovcov a je spojený s urýchlením ich vývoja skoré štádia embryogenéza. O U primitívnejších strunatcov, ako je lancelet, sa chordomesoderm zvyčajne tvorí na začiatku ďalšej fázy po gastrulácii – organogenéze. Posun v čase vývoja niektorých orgánov voči iným u potomkov v porovnaní s predkami je prejavom heterochrónie. Zmeniťčas uloženia najdôležitejších orgánov v procese evolúcie nie je nezvyčajný.
Gastrulačnú fázu charakterizujú dôležité bunkové transformácie, ako sú smerové pohyby skupín a jednotlivých buniek, selektívne rozmnožovanie a triedenie buniek, začiatok cytodiferenciácie a indukčné interakcie.
Gastrulačné metódy
Rozlišujú sa štyri typy priestorovo usmernených pohybov buniek, ktoré vedú k premene embrya z jednej vrstvy na viacvrstvovú.
- Intususcepcia – invaginácia jedného z úsekov blastodermu dovnútra ako celá vrstva. V lancelete invaginujú bunky vegetatívneho pólu, u obojživelníkov dochádza k intususcepcii na hranici medzi zvieracím a vegetatívnym pólom v oblasti sivého polmesiaca. Proces invaginácie je možný iba vo vajciach s malým alebo stredným množstvom žĺtka.
- epiboly – zanášanie malými bunkami živočíšneho pólu väčších, zaostávajúcich v rýchlosti delenia a menej pohyblivých buniek vegetatívneho pólu. Tento proces je jasne vyjadrený u obojživelníkov.
- Denominácia – stratifikácia blastodermových buniek do dvoch vrstiev ležiacich nad sebou. Delamináciu možno pozorovať v diskoblastule embryí s čiastočným typom drvenia, ako sú plazy, vtáky a vajcorodé cicavce. Delaminácia sa prejavuje v embryoblastoch placentárnych cicavcov, čo vedie k vytvoreniu hypoblastu a epiblastu.
- imigrácia – pohyb skupín alebo jednotlivých buniek, ktoré nie sú spojené do jednej vrstvy. Imigrácia sa vyskytuje u všetkých embryí, ale je najcharakteristickejšia pre druhú fázu gastrulácie u vyšších stavovcov.
Morfológia gastrulácie

Invaginácia začína na vegetatívnom póle. Vďaka rýchlejšiemu deleniu bunky živočíšneho pólu rastú a vytláčajú bunky vegetatívneho pólu do blastuly. To je uľahčené zmenou stavu cytoplazmy v bunkách, ktoré tvoria pery blastopóru a priľahlé k nim. V dôsledku invaginácie klesá blastocoel a zvyšuje sa gastrocoel. Súčasne s vymiznutím blastocoelu sa ektoderm a endoderm dostanú do úzkeho kontaktu. V lancelete, ako vo všetkých deuterostómoch (zahŕňajú typ ostnatokožca, typ strunatca a niektoré ďalšie malé druhy živočíchov), sa oblasť blastopóru mení na chvostovú časť organizmu, na rozdiel od protostómov, v ktorých blastopor zodpovedá do hlavovej časti. Ústny otvor v deuterostómoch sa tvorí na konci embrya oproti blastopóru.
Gastrulácia u obojživelníkov má veľa spoločného s gastruláciou lanceletu, ale keďže žĺtok v ich vajciach je oveľa väčší a nachádza sa hlavne na vegetatívnom póle, veľké blastoméry amfiblastuly sa nedokážu vydúvať dovnútra. Intususcepcia je trochu iná. Na hranici medzi zvieracím a vegetatívnym pólom v oblasti sivého kosáka sa bunky najskôr silne natiahnu dovnútra, pričom majú formu „baňovitého tvaru“ a potom so sebou ťahajú bunky povrchovej vrstvy blastuly. Objaví sa polmesiačiková ryha a dorzálny blastopórový pysk.
Súčasne sa menšie bunky zvieracieho pólu, ktoré sa delia rýchlejšie, začnú pohybovať smerom k vegetatívnemu pólu. V oblasti chrbtovej pery sa vytáčajú a invaginujú a väčšie bunky rastú po stranách a na opačnej strane kosákovitej ryhy. Proces epiboly potom vedie k vytvoreniu laterálnych a ventrálnych pyskov blastopóru. Blatopór sa uzatvára do prstenca, vo vnútri ktorého sú nejaký čas viditeľné veľké svetelné bunky vegetatívneho pólu v podobe takzvanej žĺtkovej zátky. Neskôr sú úplne ponorené dovnútra a blastopór sa zužuje.
Metódou značenia vitálnymi (vitálnymi) farbivami u obojživelníkov boli podrobne študované pohyby buniek blastuly počas gastrulácie.Zistilo sa, že špecifické oblasti blastodermy, nazývané presumptívne, sú počas normálneho vývoja najskôr v zložení určitých rudimentov orgánov a potom v zložení samotných orgánov.
Analýza skorých štádií vývoja obojživelníkov nám umožňuje dospieť k záveru, že ovoplazmatická segregácia, ktorá sa jasne prejavuje vo vajíčku a zygote, má veľký význam pri určovaní osudu buniek, ktoré zdedili určitú časť cytoplazmy. Určitá podobnosť medzi procesmi gastrulácie a oblasťou predpokladaných orgánov u obojživelníkov a lanceletu, t.j. homológia hlavných orgánov, ako je nervová trubica, notochorda, sekundárne črevo, naznačuje ich fylogenetický vzťah.
Gastrulácia v embryách s meroblastickým typom štiepenia (čiastočné delenie vajíčka) a vývoj má svoje vlastné charakteristiky. U vtákov začína po rozdrvení a vytvorení blastuly počas prechodu embrya cez vajcovod. V čase kladenia vajíčka sa embryo už skladá z niekoľkých vrstiev: horná vrstva sa nazýva epiblast, spodná vrstva sa nazýva primárny hypoblast. Medzi nimi je úzka medzera - blastocoel. Potom sa vytvorí sekundárny hypoblast, ktorého spôsob vzniku nie je celkom jasný. Existujú dôkazy, že primárne zárodočné bunky pochádzajú z primárneho hypoblastu vtákov, zatiaľ čo sekundárne tvorí extraembryonálny endoderm. Vznik primárneho a sekundárneho hypoblastu sa považuje za jav predchádzajúci gastrulácii.
Hlavné udalosti gastrulácie a konečnej tvorby troch zárodočných vrstiev sa začínajú po kladení vajíčok so začiatkom inkubácie. Dochádza k hromadeniu buniek v zadnej časti epiblastu v dôsledku nerovnomernej rýchlosti bunkového delenia a ich pohybu z laterálnych častí epiblastu do stredu, smerom k sebe. Vytvorí sa takzvaný primárny pruh, ktorý sa tiahne smerom k hlavovému koncu. Primárna drážka je vytvorená v strede primárneho pásu a primárne hrebene sú vytvorené pozdĺž okrajov. Na hlavovom konci primárneho pruhu sa objaví zhrubnutie - Hensenov uzol a v ňom - primárna jamka.
Keď bunky epiblastu vstúpia do primárnej drážky, ich tvar sa zmení. Tvarom sa podobajú na bunky „baňovitého tvaru“ gastruly obojživelníkov. Tieto bunky sa potom stanú hviezdicovitými a ponoria sa pod epiblast, aby vytvorili mezoderm. Endoderm sa tvorí na báze primárneho a sekundárneho hypoblastu s pridaním novej generácie endodermálnych buniek migrujúcich z horných vrstiev, blastodermy. Prítomnosť niekoľkých generácií endodermálnych buniek naznačuje predĺženie doby gastrulácie v čase.
Časť buniek migrujúcich z epiblastu cez Hensenov uzol tvorí budúci notochord. Súčasne s iniciáciou a predĺžením chordy postupne mizne Hensenov uzol a primárny pruh v smere od predného ku kaudálnemu koncu. Tomu zodpovedá zúženie a uzavretie blastopóru. Keď sa primárny pruh zmršťuje, zanecháva za sebou vytvorené časti osových orgánov embrya v smere od hlavy k častiam chvosta. Zdá sa rozumné považovať pohyby buniek v embryu kurčiat za homológne epiboly a primárny pruh a Hensenov uzol za homológne s blastopórom v dorzálnej pere gastruly obojživelníka.
Vlastnosti štádia gastrulácie
Gastrulácia je charakterizovaná rôznymi bunkovými procesmi. Mitotická reprodukcia buniek pokračuje a má rôznu intenzitu v rôznych častiach embrya. Najcharakteristickejším znakom gastrulácie je však pohyb bunkových hmôt. To vedie k zmene štruktúry embrya a jeho premene z blastuly na gastrulu. Bunky sú triedené podľa ich príslušnosti k rôznym zárodočným vrstvám, vo vnútri ktorých sa navzájom „rozoznávajú“.
Gastrulačnou fázou sa začína cytodiferenciácia, ktorá znamená prechod k aktívnemu využívaniu biologickej informácie vlastného genómu. Jeden z regulátorov genetickej aktivity je rôzny chemické zloženie cytoplazma embryonálnych buniek, vytvorená ako výsledok ovoplazmatickej segregácie. Takže ektodermálne bunky obojživelníkov majú tmavú farbu kvôli pigmentu, ktorý sa do nich dostal zo zvieracieho pólu vajíčka, a endodermálne bunky sú svetlé, pretože pochádzajú z vegetatívneho pólu vajíčka.
Počas gastrulácie je úloha embryonálnej indukcie veľmi veľká. Ukázalo sa, že výskyt primárneho pruhu u vtákov je výsledkom indukčnej interakcie medzi hypoblastom a epiblastom. Hypoblast má polaritu. Zmena polohy hypoblastu voči epiblastu spôsobuje zmenu orientácie primitívneho pruhu.
Všetky tieto procesy sú podrobne popísané v kapitole 8.2. Treba poznamenať, že také prejavy integrity embrya, ako je determinácia, embryonálna regulácia a integrácia, sú mu vlastné počas gastrulácie v rovnakej miere ako počas drvenia.
Histogenéza + organogenéza

Organogenéza, ktorá spočíva vo vytváraní jednotlivých orgánov, tvorí hlavnú náplň embryonálneho obdobia. Pokračujú v larve a končia v juvenilnom období. Organogenéza sa vyznačuje najkomplexnejšími a najrozmanitejšími morfogenetickými premenami. Nevyhnutným predpokladom prechodu k organogenéze je dosiahnutie štádia gastruly zárodkom, a to vytvorenie zárodočných vrstiev. Zárodočné vrstvy, ktoré vo vzťahu k sebe zaujímajú určitú pozíciu, poskytujú kontaktom a interakciou také vzťahy medzi rôznymi bunkovými skupinami, ktoré stimulujú ich vývoj v určitom smere. Táto takzvaná embryonálna indukcia je najdôležitejším dôsledkom interakcie medzi zárodočnými vrstvami.
V priebehu organogenézy sa mení tvar, štruktúra a chemické zloženie buniek, izolujú sa bunkové skupiny, ktoré sú základom budúcich orgánov. Postupne sa vyvíja určitá forma orgánov, vytvárajú sa medzi nimi priestorové a funkčné spojenia. Procesy morfogenézy sú sprevádzané diferenciáciou tkanív a buniek, ako aj selektívnym a nerovnomerným rastom jednotlivých orgánov a častí tela. Predpokladom organogenézy spolu s reprodukciou buniek, migráciou a triedením je ich selektívna smrť.
Samotný začiatok organogenézy sa nazýva neurulácia. Neurulácia pokrýva procesy od objavenia sa prvých príznakov tvorby nervovej platničky až po jej uzavretie v nervovej trubici. Paralelne sa vytvára notochord a sekundárne črevo a mezoderm ležiaci po stranách notochordu sa v kraniokaudálnom smere rozdeľuje na segmentované párové štruktúry - somity.
Nervový systém stavovcov vrátane človeka sa vyznačuje stabilitou hlavného štrukturálneho plánu počas evolučnej histórie podtypu. Pri tvorbe neurálnej trubice majú všetky strunatce veľa spoločného. Spočiatku sa nešpecializovaný dorzálny ektoderm, ktorý reaguje na indukčný účinok z chordomesodermu, zmení na nervovú platňu reprezentovanú cylindrickými neuroepitelovými bunkami.
Nervová platnička nezostáva dlho sploštená. Čoskoro sa jeho bočné okraje zdvihnú a vytvoria nervové záhyby, ktoré ležia na oboch stranách plytkej pozdĺžnej nervovej drážky. Okraje nervových záhybov sa potom uzavrú a vytvoria uzavretú nervovú trubicu s kanálom vo vnútri - neurocoel. V prvom rade k uzavretiu nervových záhybov dochádza na úrovni začiatku miechy a potom sa šíri v hlave a smery chvosta. Ukázalo sa, že mikrotubuly a mikrofilamenty neuroepiteliálnych buniek hrajú dôležitú úlohu v morfogenéze nervovej trubice. Deštrukcia týchto bunkových štruktúr kolchicínom a cytochalazínom B spôsobuje, že nervová platnička zostáva otvorená. Neuzavretie nervových záhybov vedie k vrodeným malformáciám nervovej trubice.
Po uzavretí nervových záhybov tvoria bunky, ktoré sa pôvodne nachádzali medzi nervovou platničkou a budúcim kožným ektodermom, nervový hrebeň. Bunky neurálnej lišty sa vyznačujú schopnosťou extenzívne migrovať, ale vysoko regulovaným spôsobom po celom tele a tvoria dva hlavné prúdy. Bunky jedného z nich - povrchové - sú zahrnuté v epidermis alebo dermis kože, kde sa diferencujú na pigmentové bunky. Ďalší prúd migruje brušným smerom, vytvára senzitívne miechové gangliá, sympatické gangliá, dreň nadobličiek, parasympatické gangliá. Bunky z lebečnej neurálnej lišty vedú k vzniku nervových buniek a mnohých ďalších štruktúr, ako je žiabrová chrupavka, niektoré pokrývajúce kosti lebky.
Mezoderm, ktorý zaberá miesto po stranách notochordu a siaha ďalej medzi kožným ektodermom a endodermou sekundárneho čreva, sa delí na dorzálnu a ventrálnu oblasť. Dorzálna časť je segmentovaná a reprezentovaná párovými somitmi. Kladenie somitov prebieha od hlavy po chvost. Ventrálna časť mezodermu, ktorá vyzerá ako tenká vrstva buniek, sa nazýva laterálna platňa. Somity sú spojené s laterálnou platňou intermediárnym mezodermom vo forme segmentovaných nôh somitu.
Všetky oblasti mezodermu sa postupne diferencujú. Na začiatku tvorby majú somity konfiguráciu charakteristickú pre epitel s dutinou vo vnútri. Pod indukčným pôsobením vychádzajúcim z notochordu a neurálnej trubice sa ventromediálne časti somitov - sklerotómy - menia na sekundárny mezenchým, sú zo somitu vypudené a obklopujú notochord a ventrálnu časť neurálnej trubice. Nakoniec sa z nich vytvoria stavce, rebrá a lopatky.
Dorzolaterálna časť somitov s vnútri tvorí myotómy, z ktorých sa vyvinie priečne pruhované kostrové svalstvo tela a končatín. Vonkajšia dorzolaterálna časť somitov tvorí dermatómy, z ktorých vzniká vnútorná vrstva kože – dermis. Z oblasti nôh somitov so základmi nefrota a gonotu sa tvoria vylučovacie orgány a pohlavné žľazy.
Pravá a ľavá nesegmentovaná bočná platnička sa rozdelí na dva pláty, čím je ohraničená sekundárna telová dutina – celok. Vnútorný list susediaci s endodermom sa nazýva viscerálny. Obklopuje črevo zo všetkých strán a tvorí mezentérium, pokrýva pľúcny parenchým a srdcový sval. Vonkajšia vrstva laterálnej dosky susedí s ektodermou a nazýva sa parietálna. V budúcnosti tvorí vonkajšie listy pobrušnice, pohrudnice a osrdcovníka.
Endoderm vo všetkých embryách nakoniec tvorí epitel sekundárneho čreva a mnohých jeho derivátov. Samotné sekundárne črevo sa vždy nachádza pod akordom.
V procese neurulácie tak vzniká komplex axiálnych orgánov - nervová trubica - notochorda - črevo, ktoré sú najcharakteristickejším znakom organizácie tela všetkých strunatcov. Rovnaký pôvod, vývoj a vzájomné usporiadanie osových orgánov prezrádza ich úplnú homológiu a evolučnú kontinuitu.
Hĺbkové skúmanie a porovnanie neurulačných procesov u konkrétnych predstaviteľov strunatcového typu odhaľuje niektoré rozdiely, ktoré sú spojené najmä so znakmi, ktoré závisia od štruktúry vajíčok, spôsobu drvenia a gastrulácie. Pozornosť upriamuje na odlišný tvar embryí a posun v čase kladenia osových orgánov voči sebe, t.j. heterochrónia opísaná vyššie.
Ektoderm, mezoderm a endoderm sa v priebehu ďalšieho vývoja, ktoré sa navzájom ovplyvňujú, podieľajú na tvorbe určitých orgánov. Vznik rudimentu orgánu je spojený s miestnymi zmenami v určitej oblasti zodpovedajúcej zárodočnej vrstvy. Takže z ektodermy sa vyvíja epidermis kože a jej deriváty (perie, vlasy, nechty, koža a mliečne žľazy), zložky orgánov videnia; sluch, čuch, epitel ústnej dutiny, zubná sklovina. Najdôležitejšími ektodermálnymi derivátmi sú nervová trubica, neurálna lišta a všetky nervové bunky z nich vytvorené.
Deriváty endodermu sú epitel žalúdka a čriev, pečeňové bunky, sekrečné bunky pankreasu, črevné a žalúdočné žľazy. Predná časť embryonálneho čreva tvorí epitel pľúc a dýchacích ciest, ako aj secernujúce bunky predného a stredného laloka hypofýzy, štítnej žľazy a prištítnych teliesok.
Mezoderm, okrem už vyššie popísaných kostrových štruktúr, kostrové svaly, dermis kože, orgány vylučovacej a reprodukčnej sústavy, tvorí kardiovaskulárny systém, lymfatický systém, pohrudnicu, pobrušnicu a osrdcovník. Z mezenchýmu, ktorý má zmiešaný pôvod vďaka bunkám troch zárodočných vrstiev, sa vyvíjajú všetky druhy väziva, hladké svaly, krv a lymfa.
Rudiment konkrétneho orgánu sa spočiatku tvorí zo špecifickej zárodočnej vrstvy, ale potom sa orgán stáva zložitejším a v dôsledku toho sa na jeho tvorbe podieľajú dve alebo tri zárodočné vrstvy.
Periodizácia ľudskej embryogenézy

V ľudskej embryogenéze existujú 4 obdobia:
- Počiatočné (1 týždeň vývoja, až do implantácie embrya do sliznice maternice).
- Embryonálne (2-8 týždňov).
- Prefetálny (9-12 týždňov) = larva u zvierat
- Fetálny (13 týždňov - pôrod) = metamorfóza
V embryonálnom období dochádza k gastrulácii, blastulácii a neurulácii. V prefetálnom období prebieha intenzívna organogenéza, anatomické uloženie orgánov. Fetálne obdobie je charakterizované tvorbou plodu pod ochranou membrán.
V počiatočnom období je zygota- 1 embryonálna bunka, v nej sa určujú samostatné úseky cytoplazmy, dochádza k syntéze DNA a bielkovín.
Štádium štiepenia je obdobím intenzívneho delenia buniek. Veľkosť embrya sa nezvyšuje a syntetické procesy sú aktívne. Dochádza k intenzívnej syntéze DNA, RNA, histónu a iných proteínov.
| Vývojové obdobie, týždne |
Morfogenetické procesy |
| Počiatočné obdobie (skorá embryogenéza) | |
| 1 |
Hnojenie. Štiepenie zygoty. Tvorba moruly a blastuly. Prvá fáza gastrulácie (delaminácia), tvorba epiblastu a hypoblastu. Začiatok implantácie. |
| Embryonálne obdobie (embryonálne) | |
| 2 |
Dokončenie implantácie. Tvorba zárodočného disku. Druhá etapa gastrulácie (imigrácia), tvorba primárneho pruhu, prechordálnej platničky. Tvorba amniotických a germinálnych vezikúl, extraembryonálny mezoderm. Diferenciácia trofoblastu na cytotrofoblast a symplastotropoblast, primárne choriové klky. Vývoj primárneho a sekundárneho (definitívneho) žĺtkového vaku. |
| 3 |
Pokračovanie 2. štádia gastrulácie, tvorba troch zárodočných vrstiev, notochorda, prechordálna platnička, neurálna trubica, neurálna lišta. Začiatok segmentácie dorzálneho mezodermu (somity, segmentové nohy), tvorba parietálnych a viscerálnych listov splanchnotómov a embryonálneho coelomu, ktorý sa ďalej delí na tri telové dutiny – perikardiálnu, pleurálnu, peritoneálnu. Položenie srdca, ciev, pronephros - pronephros. Tvorba extraembryonálnych orgánov - alantois, sekundárne a terciárne choriové klky. Tvorba kmeňového záhybu a oddelenie primárneho čreva embrya od sekundárneho žĺtkového vaku. |
| 4 |
Prehĺbenie žĺtkového záhybu, tvorba stopky žĺtka a elevácia embrya v amniovej dutine. Pokračovanie segmentácie dorzálneho mezodermu do 30 somitov a diferenciácia na myotóm, sklerotóm a dermatóm. Uzavretie nervovej trubice a tvorba predného neuropóru (do 25. dňa) a zadného neuropóru (do 27. dňa), tvorba nervových ganglií; uloženie pľúc, žalúdka, pečene, pankreasu, endokrinných žliaz (adenohypofýza, štítna žľaza a prištítne telieska). Tvorba plaku ucha a šošovky, primárna oblička - mezonefros. Začiatok tvorby placenty. Tvorba rudimentov horných a dolných končatín, 4 páry žiabrových oblúkov. |
| 5 |
Rozšírenie predného konca nervovej trubice. Koniec segmentácie mezodermu (tvorba 42-44 párov somitov), tvorba nesegmentovaného mezodermu (nefrogénneho tkaniva) v kaudálnej oblasti. Vývoj priedušiek a lalokov pľúc. Položenie poslednej obličky (metanefros), urogenitálny sínus, konečník, močového mechúra. Tvorba genitálnych hrebeňov. |
| 6 |
Formovanie tváre, prstov. Začiatok tvorby vonkajšieho ucha a očnej gule. Tvorba základov mozgu - most, cerebellum. Tvorba pečene, pankreasu, pľúc. Ukladanie mliečnych žliaz. Oddelenie pohlavných žliaz od mezonefrosu, vznik pohlavných rozdielov v pohlavných žľazách. |
| 7 |
Formovanie horných a dolných končatín. Roztrhnutie kloakálnej membrány. |
| 8 |
Tvorba prstov horných a dolných končatín. Výrazné zvýšenie veľkosti hlavy (až o 1/2 dĺžky tela). Pupočná šnúra. |
| plodné obdobie | |
| 9 |
Dokončenie tvorby placenty (12 - 13 týždňov). Tvorba hladkého a vilózneho chorionu. Rast symplastotropoblastu a redukcia cytotrofoblastu v placentárnych klkoch. Výrazné zvýšenie veľkosti a hmotnosti plodu. Pokračovanie procesov tvorby tkanív a orgánov. Tvorba systému matka-plod. Fetálny obeh. |
Kritické obdobia D Y

Kritické obdobia- obdobia, v ktorých existujú spoločné a špecifické znaky v povahe reakcií embrya a plodu na patogénne účinky. Vyznačujú sa prevahou procesov aktívnej bunkovej a tkanivovej diferenciácie a výrazným zvýšením metabolických procesov.
- 1 kritické obdobie od 0 do 8 dní. Zvažuje sa od okamihu oplodnenia vajíčka až po zavedenie blatocysty do decidua. Počas tohto obdobia neexistuje spojenie medzi embryom a telom matky. Poškodzujúce faktory buď nespôsobia smrť plodu, alebo embryo zomrie (princíp „všetko alebo nič“). charakteristický znak obdobie je absencia malformácií aj pod vplyvom environmentálnych faktorov, ktoré majú výrazný teratogénny účinok. Výživa embrya je autotropná – vďaka látkam obsiahnutým vo vajíčku, a potom vďaka tekutej sekrécii trofoblastu v dutine blastocysty.
- 2. kritické obdobie od 8 dní do 8 týždňov. Počas tohto obdobia dochádza k tvorbe orgánov a systémov, v dôsledku čoho je charakteristický výskyt viacerých malformácií. Najcitlivejšia fáza je prvých 6 týždňov: sú možné poruchy centrálneho nervového systému, sluchu a očí. Pod vplyvom poškodzujúcich faktorov dochádza spočiatku k inhibícii a zastaveniu vývoja, potom k náhodnému množeniu niektorých a dystrofii iných rudimentov orgánov a tkanív. Hodnota poškodenia nie je ani tak gestačný vek, ale trvanie vystavenia nepriaznivému faktoru.
- 3. kritické obdobie - 3-8 týždňov vývoja. Spolu s organogenézou dochádza k tvorbe placenty a chorionu. Pri vystavení škodlivému faktoru je narušený vývoj alantoisu, ktorý je veľmi citlivý na poškodenie: dochádza k vaskulárnej smrti, v dôsledku ktorej sa zastaví vaskularizácia chorionu s nástupom primárnej placentárnej nedostatočnosti.
- 4. kritické obdobie - 12.-14. Vzťahuje sa na vývoj plodu. Nebezpečenstvo je spojené s tvorbou vonkajších genitálií u ženských plodov s tvorbou falošného mužského hermafroditizmu.
- 5. kritické obdobie - 18-22 týždňov. V tomto období sa dokončuje tvorba nervového systému, zaznamenáva sa bioelektrická aktivita mozgu, zmeny krvotvorby, tvorba niektorých hormónov.
VÝVOJ EMBRYA
Podstata fázy drvenia. Rozdelenie - ide o sériu postupných mitotických delení zygoty a ďalších blastomér, ktoré končia vytvorením mnohobunkového embrya - blastula. Prvé rozdelenie štiepenia začína po spojení dedičného materiálu pronuklea a vytvorení spoločnej metafázovej platničky. Bunky vzniknuté pri štiepení sa nazývajú blastoméry(z gréčtiny. výbuch- klíčok, klíčok). Znakom mitotických delení drvenia je, že s každým delením sa bunky zmenšujú a zmenšujú, až kým nedosiahnu pomer objemov jadra a cytoplazmy, ktorý je obvyklý pre somatické bunky. Napríklad u morského ježka to vyžaduje šesť delení a embryo pozostáva zo 64 buniek. Medzi po sebe nasledujúcimi deleniami nedochádza k rastu buniek, ale DNA sa nevyhnutne syntetizuje.
Všetky prekurzory DNA a potrebné enzýmy sa akumulujú počas oogenézy. V dôsledku toho sa mitotické cykly skracujú a delenia nasledujú po sebe oveľa rýchlejšie ako v bežných somatických bunkách. Po prvé, blastoméry spolu susedia a vytvárajú zhluk buniek tzv morula. Potom sa medzi bunkami vytvorí dutina - blastocoel, naplnené kvapalinou. Bunky sú vytlačené na perifériu a tvoria stenu blastuly - blastoderm. Celková veľkosť embrya na konci štiepenia v štádiu blastuly nepresahuje veľkosť zygoty.
Hlavným výsledkom obdobia drvenia je premena zygoty na mnohobunkové jednozmenné embryo.
Morfológia drvenia. Spravidla sú blastoméry usporiadané v prísnom poradí voči sebe navzájom a voči polárnej osi vajíčka. Poradie alebo spôsob drvenia závisí od množstva, hustoty a rozloženia žĺtka vo vajci. Podľa pravidiel Sachs-Hertwigovej má bunkové jadro tendenciu byť umiestnené v strede cytoplazmy bez žĺtka a vreteno bunkového delenia - v smere najväčšieho rozsahu tejto zóny.
V oligo- a mesolecitálnych vajíčkach štiepenie kompletný, alebo holoblastický. Tento typ drvenia sa vyskytuje u mihule, niektorých rýb, všetkých obojživelníkov, ako aj u vačkovcov a placentárnych cicavcov. Pri úplnom delení rovina prvého delenia zodpovedá rovine bilaterálnej symetrie. Rovina druhého delenia prebieha kolmo na rovinu prvého. Obe brázdy prvých dvoch divízií sú poludníky, ᴛ.ᴇ. začínajú na zvieracom póle a šíria sa k vegetatívnemu pólu. Vaječná bunka je rozdelená na štyri blastoméry, ktoré majú viac-menej rovnakú veľkosť. Rovina tretieho delenia prebieha kolmo na prvé dve v smere zemepisnej šírky. Potom sa v mezolecitálnych vajíčkach v štádiu ôsmich blastomérov prejavuje nerovnomerné drvenie. Na zvieracom póle sú štyri menšie blastoméry - mikrometre, na vegetatívne - štyri väčšie - makroméry. Potom rozdelenie opäť ide v rovinách poludníka a potom znova v rovinách zemepisnej šírky.
V polylecitálnych oocytoch kostnatých rýb, plazov, vtákov a tiež monotrémnych cicavcov štiepenie čiastočný, alebo meroblastický,ᴛ.ᴇ. pokrýva iba cytoplazmu bez žĺtka. Nachádza sa vo forme tenkého disku pri zvieracom póle, v súvislosti s tým sa tento typ drvenia nazýva tzv. diskoidný.
Pri charakterizácii typu drvenia sa berie do úvahy aj relatívna poloha a rýchlosť delenia blastomérov. Ak sú blastoméry usporiadané v radoch nad sebou pozdĺž polomerov, nazýva sa drvenie radiálne. Je typický pre strunatce a ostnatokožce. V prírode existujú iné varianty priestorového usporiadania blastomérov počas drvenia, ktoré určuje také typy, ako sú špirálové u mäkkýšov, bilaterálne u ascaris, anarchické u medúz.
Bol zaznamenaný vzťah medzi distribúciou žĺtka a stupňom synchronizácie pri delení živočíšnych a vegetatívnych blastomér. V oligolecitálnych vajíčkach ostnatokožcov je štiepenie takmer synchrónne, v bunkách mezolecitálnych vajíčok je synchronizácia narušená po treťom delení, pretože vegetatívne blastoméry sa delia pomalšie kvôli veľkému množstvu žĺtka. Vo formách s čiastočným štiepením sú delenie od samého začiatku asynchrónne a blastoméry zaujímajúce centrálnu polohu sa delia rýchlejšie.
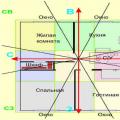
Ryža. 7.2. Štiepenie u strunatcov s rôznymi druhmi vajec.
A - lancelet; B -žaba; IN - vták; G - cicavec:
ja- dve blastoméry II-štyri blastoméry, III- osem blastomérov, IV- morula, V- blastula;
1 - drvenie brázd, 2 - blastoméry, 3- blastoderm, 4- blastoiel, 5- epiblast, 6- hypoblast, 7-embryoblast, 8- trofoblast; veľkosti jadier na obrázku neodrážajú skutočné pomery veľkosti
Ryža. 7.2. Pokračovanie
Na konci drvenia sa vytvorí blastula. Typ blastuly závisí od typu drvenia, a teda od typu vajíčka. Niektoré typy drvenia a blastuly sú znázornené na obr. 7.2 a schémy 7.1. Viac Detailný popis drvenie u cicavcov a ľudí, pozri ods. 7.6.1.
Vlastnosti molekulárno-genetických a biochemických procesov pri drvení. Ako je uvedené vyššie, mitotické cykly počas obdobia štiepenia sú značne skrátené, najmä na samom začiatku.
Napríklad celý cyklus štiepenia u vajíčok morského ježka trvá 30-40 minút, pričom trvanie S-fázy je len 15 minút. gi- a 02-periódy prakticky chýbajú, keďže potrebná zásoba všetkých látok sa vytvorila v cytoplazme vaječnej bunky a čím je väčšia, tým je väčšia. Pred každým delením dochádza k syntéze DNA a histónov.
Rýchlosť, ktorou sa replikačná vidlica pohybuje pozdĺž DNA počas štiepenia, je normálna. Zároveň je v DNA blastomér viac iniciačných bodov ako v somatických bunkách. Syntéza DNA prebieha vo všetkých replikónoch súčasne, synchrónne. Z tohto dôvodu sa čas replikácie DNA v jadre zhoduje s časom zdvojnásobenia jedného, navyše skráteného, replikónu. Ukázalo sa, že pri odstránení jadra zo zygoty dochádza k štiepeniu a embryo vo svojom vývoji dosiahne takmer štádium blastuly. Ďalší vývoj sa zastaví.
Na začiatku štiepenia prakticky chýbajú iné typy jadrovej aktivity, ako je transkripcia. IN odlišné typy Vo vajíčkach začína transkripcia génov a syntéza RNA v rôznych štádiách. Keď je v cytoplazme veľa rôzne látky, ako napríklad u obojživelníkov sa transkripcia neaktivuje okamžite. Syntéza RNA v nich začína v štádiu ranej blastuly. Naopak, u cicavcov sa syntéza RNA začína už v štádiu dvoch blastomér.
Počas obdobia štiepenia sa tvorí RNA a proteíny, podobné tým, ktoré sa syntetizujú počas oogenézy. Ide najmä o históny, proteíny bunkovej membrány a enzýmy potrebné na delenie buniek. Tieto proteíny sa používajú okamžite spolu s proteínmi uloženými skôr v cytoplazme oocytov. Spolu s tým je počas obdobia drvenia možná syntéza bielkovín, ktorá tam predtým nebola. Podporujú to údaje o prítomnosti regionálnych rozdielov v syntéze RNA a proteínov medzi blastomérmi. Niekedy tieto RNA a proteíny vstupujú do činnosti v neskoršom štádiu.
Dôležitú úlohu pri fragmentácii hrá delenie cytoplazmy - cytotómiu. Má osobitný morfogenetický význam, pretože určuje typ drvenia. V procese cytotómie sa najskôr vytvorí zúženie pomocou kontraktilného prstenca mikrofilamentov. Zostavenie tohto prstenca prebieha pod priamym vplyvom pólov mitotického vretienka. Po cytotómii zostávajú blastoméry oligolecitálnych vajíčok navzájom spojené len tenkými mostíkmi. Práve v tomto čase sa dajú najľahšie oddeliť. Je to preto, že cytotómia vedie k zníženiu oblasti kontaktu medzi bunkami v dôsledku obmedzeného povrchu membrán.
Ihneď po cytotómii začína syntéza nových častí bunkového povrchu, kontaktná zóna sa zvyšuje a blastoméry sa začínajú tesne dotýkať. Po hraniciach medzi jednotlivými úsekmi ovoplazmy prebiehajú štiepne brázdy, ktoré odrážajú fenomén ovoplazmatickej segregácie. Z tohto dôvodu sa cytoplazma rôznych blastomérov líši v chemickom zložení.
Drvenie - pojem a druhy. Klasifikácia a vlastnosti kategórie "Drvenie" 2017, 2018.
Na konci XII - začiatku XIII storočia. na základe všeobecného sociálneho a hospodárskeho obrodenia Nemecka došlo k dôležitým zmenám v politickej štruktúre ríše: bývalé feudálne oblasti (vojvodstvá, arcibiskupstvá) sa zmenili na takmer úplne samostatné štáty .... .
HNOJENIE Prednáška 8 Oplodnenie je stimulácia vajíčka spermiou, aby sa vyvinula, pričom sa do vajíčka súčasne prenáša dedičný materiál otca. V procese oplodnenia sa spermie spoja s vajíčkom, zatiaľ čo haploidné jadro ....
PRÍPRAVNÉ PROCESY PREDNÁŠKA č. 4 Premývanie nerastov Premývanie sa používa pri obohacovaní aluviálnych ložísk vzácnych a drahých kovov, rúd železných kovov, fosforitov, kaolínov, stavebných materiálov (piesok, drvený kameň), ...
Embryonálny vývoj je zložitý a zdĺhavý morfogenetický proces, pri ktorom z otcovských a materských zárodočných buniek vzniká nový mnohobunkový organizmus schopný samostatného života v prostredí. Je základom pohlavného rozmnožovania a zabezpečuje prenos dedičných vlastností z rodičov na potomkov.
Hnojenie spočíva v spojení spermie s vajíčkom Najdôležitejšie fázy procesu oplodnenia sú:
1) prienik spoločného podniku do vajíčka;
2) aktivácia rôznych syntetických procesov vo vajci;
3) fúzia jadier vajíčka a SP s obnovením diploidnej sady chromozómov.
Aby mohlo dôjsť k oplodneniu, je potrebná konvergencia ženských a mužských zárodočných buniek. Dosahuje sa insemináciou.
Prienik SP do vajíčka uľahčuje enzým hyaluronidáza a ďalšie biologicky aktívne látky (spermolyzín), ktoré zvyšujú permeabilitu hlavnej medzibunkovej látky Enzýmy sú vylučované akrozómom v procese akrozómová reakcia. Jeho podstata je nasledovná, v momente kontaktu s vajíčkom v hornej časti hlavičky spermie sa plazmatická membrána a priľahlá membrána rozpustia a priľahlá časť vaječnej membrány sa rozpustí. Akrozomálna membrána vyčnieva smerom von a vytvára sa výrastok vo forme dutej trubice.alebo tuberkula oplodnenia.Potom sa plazmatické membrány oboch gamét spoja a začne sa spojenie ich obsahu.Od tohto momentu sú SP a I jedna bunka zygoty.
Aktivácia I resp kortikálna reakcia, ktorý vzniká v dôsledku kontaktu so SP, má morfologické a biochemické prejavy. Prejavom aktivácie sú zmeny v povrchovej kortikálnej vrstve ooplazmy a útvaru oplodňovacie membrány. Hnojivá membrána chráni yatsyo pred prenikaním nadmerne veľkých spermií.
Zygota- diploidná (obsahujúca kompletnú dvojitú sadu chromozómov) bunka, ktorá je výsledkom oplodnenia (fúzie vajíčka a spermie). Zygota je totipotentný(teda schopný dať vzniknúť akejkoľvek inej) bunke.
U ľudí nastáva prvé mitotické delenie zygoty približne 30 hodín po oplodnení v dôsledku zložitých procesov prípravy na prvé delenie. drvenie.
Rozdelenie - je to séria postupných mitotických delení zygoty a končiaca vytvorením mnohobunkového embrya - blastula. Prvé rozdelenie štiepenia začína po spojení dedičného materiálu pronuklea a vytvorení spoločnej metafázovej platničky. Bunky vzniknuté pri štiepení sa nazývajú blastoméry(z gréčtiny. výbuch- klíčok, klíčok). Znakom delenia mitotickým štiepením je, že pri každom delení sa bunky zmenšujú a zmenšujú, až kým nedosiahnu pomer objemov jadra a cytoplazmy, ktorý je obvyklý pre somatické bunky.Po prvé, blastoméry na seba nadväzujú a vytvárajú zhluk bunky tzv morula . Potom sa medzi bunkami vytvorí dutina - blastocoel , naplnené kvapalinou. Bunky sú vytlačené na perifériu a tvoria stenu blastuly - blastoderm. Celková veľkosť embrya na konci štiepenia v štádiu blastuly nepresahuje veľkosť zygoty.
Progenéza - gametogenéza (spermato- a ovogenéza) a oplodnenie.Spermatogenéza prebieha v stočených tubuloch semenníkov a je rozdelená do štyroch období: 1) reprodukčné obdobie - I; 2) obdobie rastu - II; 3) obdobie zrenia - III; 4) obdobie formovania - IV. Ovogenéza sa uskutočňuje vo vaječníkoch a delí sa na tri obdobia: 1) reprodukčné obdobie (v embryogenéze a počas 1. roku postembryonálneho vývoja); 2) obdobie rastu (malého a veľkého); 3) obdobie dozrievania.Vajíčko pozostáva z jadra s haploidnou sadou chromozómov a výraznou cytoplazmou, ktorá obsahuje všetky organely s výnimkou cytocentra.
Rozdelenie. drviace vlastnosti. Hlavné druhy vajec podľa umiestnenia žĺtka. Vzťah štruktúry vajíčka s typom drvenia. Blastoméry a embryonálne bunky. Štruktúra a typy blastuly.
Rozdelenie - Tento proces je založený na delení mitotických buniek. Dcérske bunky vytvorené v dôsledku delenia sa však nerozchádzajú, ale zostávajú tesne vedľa seba. V procese drvenia sa dcérske bunky postupne zmenšujú. Každé zviera sa vyznačuje určitým typom drvenia v dôsledku množstva a charakteru rozloženia žĺtka vo vajci. Žĺtok bráni drveniu, preto sa časť zygoty preťažená žĺtkom štiepi pomalšie alebo sa neštiepi vôbec.
V izolecitale, nekvalitné žĺtkom oplodnené vajíčko lancety, prvá štiepna brázda v podobe medzery začína na zvieracom póle a postupne sa rozširuje v pozdĺžnom poludníkovom smere smerom k vegetatívnemu, čím sa vajíčko rozdeľuje na 2 bunky - 2 blastoméry. Druhá brázda prebieha kolmo na prvú – tvoria sa 4 blastoméry. V dôsledku série postupných delení sa vytvárajú skupiny buniek, ktoré spolu tesne susedia. U niektorých zvierat takéto embryo pripomína moruše alebo malinu. Dostal meno morula(lat. morum - moruša) - mnohobunková guľa bez dutiny vo vnútri.
IN telolecitné vajíčka , preťažený žĺtkom - drvenie môže byť úplne jednotné alebo nerovnomerné a neúplné. Blastoméry vegetatívneho pólu v dôsledku množstva inertného žĺtka vždy zaostávajú za blastomérmi živočíšneho pólu v rýchlosti štiepenia. Úplné, ale nerovnomerné štiepenie je charakteristické pre vajíčka obojživelníkov.. U rýb, vtákov a niektorých iných zvierat je rozdrvená iba časť vajíčka umiestnená na zvieracom póle; dochádza k neúplnému diskoidnému štiepeniu. V procese drvenia sa zvyšuje počet blastomérov, ale blastoméry nedorastajú do veľkosti pôvodnej bunky, ale každým drvením sa zmenšujú. Vysvetľuje to skutočnosť, že mitotické cykly drviacej zygoty nemajú typickú medzifázu; presyntetické obdobie (G1) chýba a syntetické obdobie (S) začína už v telofáze predchádzajúcej mitózy.
Štiepenie vajíčka končí tvorbou blastula.
V polylecitálnych oocytoch kostnaté ryby, plazy, vtáky, ako aj drvenie monotrémnych cicavcov čiastočný, alebo meroblastický, tie. pokrýva iba cytoplazmu bez žĺtka. Nachádza sa vo forme tenkého disku na zvieracej tyči, preto tento typ drvenie tzv diskoidný . Pri charakterizácii typu drvenia sa berie do úvahy aj relatívna poloha a rýchlosť delenia blastomérov. Ak sú blastoméry usporiadané v radoch nad sebou pozdĺž polomerov, drvenie tzv radiálne.
Fragmentácia môže byť: deterministická a regulačná; úplné (holoblastické) alebo neúplné (meroblastické); rovnomerné (blastoméry sú viac-menej rovnakej veľkosti) a nerovnomerné (blastoméry nie sú rovnako veľké, rozlišujú sa dve až tri veľkostné skupiny, zvyčajne nazývané makro- a mikroméry)
Druhy vajec:
Množstvo žĺtka - Oligolecetal (lancelet) Mesolacetal (obojživelníky) Polylecetal (ryby, vtáky)
miesto- Izolatecetal(umiestnené difúzne, rovnomerne). Obsahujú trochu žĺtka, rovnomerne rozloženého po celej bunke. Charakteristické pre ostnatokožce, nižšie strunatce a cicavce. U cicavcov sú to aleciálne vajcia (prakticky neexistuje žĺtok)
Telolecetal(s miernym množstvom žĺtka na dolnom vegetatívnom póle)
Ostro telolecitný (s veľkým množstvom žĺtka zaberá celé vajce, okrem horného pólu. Žĺtka je veľa, sústredený na vegetatívnom póle. Rozlišujú sa 2 skupiny: stredne telolektálny (mäkkýše, obojživelníky) a ostro lecitálny ( plazy a vtáky).Cytoplazma a jadro sú sústredené na anomálnom póle .
Centrolecetal(žĺtok je trochu, ale v strede hustý). Malý žĺtok, ktorý sa nachádza v strede. charakteristické pre článkonožce
Blastomeres- bunky vznikajúce v dôsledku delenia drvených vajíčok u mnohobunkových živočíchov. Charakteristickým znakom B. je absencia rastu v období medzi deleniami, v dôsledku čoho sa pri ďalšom delení objem každej B. zníži na polovicu. S holoblastom drvenie v telolecitálnych vajíčkach B. sa líšia veľkosťou: veľké B. - makroméry, stredné - mezoméry, malé - mikroméry. Počas synchrónneho štiepenia sú delenia B. spravidla vo forme homogénne, štruktúra ich cytoplazmy je veľmi jednoduchá. Potom sa povrchové B. sploštia a vajce pokračuje do fázy drvenia - blastulácia.
Štruktúra blastuly. Ak sa vytvorí pevná guľa bez dutiny vo vnútri, potom sa takéto jadro nazýva morula. Tvorba blastuly alebo moruly závisí od vlastností cytoplazmy. Blastula sa tvorí pri dostatočnej viskozite cytoplazmy, morula - pri nízkej viskozite. Pri dostatočnej viskozite cytoplazmy si blastoméry zachovávajú zaoblený tvar a v miestach kontaktu sa len mierne splošťujú. V dôsledku toho sa medzi nimi objaví medzera, ktorá sa pri zvyšovaní drvenia naplní kvapalinou a zmení sa na blastocoel. Pri nízkej viskozite cytoplazmy nie sú blastoméry zaoblené a sú umiestnené tesne vedľa seba, nevzniká medzera a nevytvára sa dutina. Blastuly sa líšia štruktúrou a závisia od typu drvenia.